Article Text

Abstract
The International Agency for Research on Cancer has recently reassessed the carcinogenicity of the biological agents classified as ‘carcinogenic to humans’. Among the biological agents having a direct role in carcinogenesis, Epstein-Barr virus, Kaposi's sarcoma-associated herpes virus and human papillomavirus contribute to a variety of malignancies worldwide in humans including nasopharyngeal carcinoma, several types of lymphomas, genital tract carcinomas and Kaposi's sarcoma. The authors review the current knowledge on cancers that have been attributed to Epstein-Barr virus, Kaposi's sarcoma-associated herpes virus and human papillomavirus looking at the pathological classification of these cancers and description of the implicated viruses, highlighting a wide range of pathological and virological diagnostic techniques. This review also focuses on the new oncological scenario ahead, once strategies against carcinogenic infectious agents are found to be effective.
- Haematopathology
- Hodgkin's disease
- cancer research
- oncology
- microbiology
- virus
Statistics from Altmetric.com
Introduction
Approximately 15–20% of cancers worldwide have been attributed to infectious agents. The second most frequent cancer in women—cervical cancer—and about 80% of liver cancers are associated with specific viral infections and are proved to be caused by such agents. The proportion of infection-attributable cancer is higher in developing countries (26%) than in developed countries (8%), reflecting the higher prevalence of infection with the major causative agents (eg, hepatitis B virus (HBV), Helicobacter pylori, human papillomavirus (HPV) and HIV-1), and lack of screening for HPV-related cervical precancerous lesions.1
The International Agency for Research on Cancer (IARC) has recently reassessed the carcinogenicity of the biological agents classified as ‘carcinogenic to humans’ (group 1), and has identified additional tumour sites and mechanisms of carcinogenesis. IARC group 1 agents assessed by the IARC monograph working group include Epstein-Barr virus (EBV), HBV, hepatitis C virus (HCV), Kaposi's sarcoma-associated herpes virus (KSHV), HIV-1, HPV-16, human T-lymphotropic virus-1 (HTLV-1), Helicobacter pylori, Clonorchis sinensis, Opistorchis viverrini and Schistosoma haematobium.2
EBV, HPV, HTLV-1 and KSHV play a direct role in carcinogenesis-encoding oncoproteins that are able to promote cellular transformation. Other infectious agents appear to have an indirect role, either inducing a chronic inflammation (HBV, HCV and Helicobacter pylori) or mediating its effects on cancer risk by lowering host immunity to other oncogenic infections (HIV-1).1
In this paper, we review the current knowledge on cancers that have been attributed to EBV, KSHV and HPV. The focus is on the classification and multidisciplinary integrative diagnosis of these cancers. The review is organised in three parts. The first is an introductory section looking at pathological classification of infection-attributable cancers with a brief description of the implicated viruses. The second section represents a wide range of pathological and virological diagnostic techniques. By reporting current knowledge, this section focuses on practical information with emphasis on clinical application. The final section, including concluding remarks, looks at the new oncological scenario that we will probably face ahead of us once strategies against carcinogenic infectious agents are found to be effective.
EBV, KSHV, HPV and attributed cancers
Epstein-Barr virusEBV is a γ-herpesvirus that has infected >90% of the world's population. Inter-human transmission occurs by the oral route, followed by productive oropharyngeal infection; then the virus infects circulating B lymphocytes, leading to the persistence of viral DNA as an episome, thus establishing a latent infection.3 ,4
Experimental data have shown substantial differences in B lymphocytes as compared with epithelial cells concerning viral gene expression, genome amplification and genome loss.5–7 A common characteristic of the virus consists of the achievement of oncogenicity after activation of intracellular signalling pathways involved in the control of proliferation.
Cancers that have been attributed to EBV
There is sufficient evidence for a causative role of EBV in nasopharyngeal carcinoma (NPC), endemic Burkitt's lymphoma (BL), immune suppression-related lymphomas, extranodal natural killer T (NK/T)-cell lymphoma (nasal type), and a subset of Hodgkin's lymphoma (HL)2 (table 1, figure 1).
Biological agents by the International Agency for Research on Cancer monograph working group (adapted from Bouvard et al 2)

Relationship of HIV-associated lymphomas with Epstein-Barr virus and Kaposi's sarcoma-associated herpes virus/HHV8-associated lymphoproliferative disorders. Reprinted with permission from Carbone A, Cesarman E, Spina M, et al. HIV-associated lymphomas and gamma-herpesviruses. Blood 2009; 113:1213–24.12 ©The American Society of Haematology. BL, Burkitt's lymphoma; DLBCL, diffuse large B-cell lymphoma; EBV, Epstein-Barr virus; HHV-8, human herpes virus 8; KSHV, Kaposi's sarcoma-associated herpes virus; MCD, multicentric Castleman's disease; PEL, primary effusion lymphoma.
NPC is rare in most populations around the world but common in southeast Asia. According to the WHO international histological classification of tumours,8 NPCs are classified into three classes: keratinising carcinoma, differentiated non-keratinising carcinoma and undifferentiated carcinoma. Most NPCs diagnosed in the ‘high-risk’ areas belong to the last two classes.8 ,9 In undifferentiated carcinoma there is a very strong association with EBV8 ,9 (figure 2). Carcinomas with a histological similarity to NPC called lymphoepithelial-like carcinomas, can occur in multiple organ sites and have been reported most frequently in the stomach. A recent review of over 30 000 cancers of the stomach identified evidence of EBV DNA in 8% of the patients.10 EBV DNA is present within tumour cells and not in the surrounding epithelium.10

Epstein-Barr virus (EBV)-associated nasopharyngeal carcinoma. Almost all carcinoma cells exhibit nuclear positivity for EBV with a strong staining intensity. EBV positivity assessed by EBER—in situ hybridisation in formalin-fixed paraffin-embedded tissue section. Haematoxylin counterstain, original magnification ×40.
Endemic BL is a highly proliferative B-cell tumour that includes three variants: endemic (affecting children in equatorial Africa and New Guinea), sporadic (affecting children and young adults all over the world), and immunodeficiency related (primarily in association with HIV infection). EBV has been proved to be present in all cases of the endemic variant, 15–20% of cases of the sporadic variant, and 30–40% of cases of the immunodeficiency-related variant.11
EBV-associated lymphomas in the immunosuppressed AIDS patients include BL, diffuse large B-cell lymphoma with immunoblastic morphology, primary central nervous system lymphoma, KSHV positive or EBV positive primary effusion lymphoma (PEL) (figure 1), and plasmablastic lymphoma of the oral cavity type (for a recent review, see Carbone et al 12). The proportion of tumours associated with EBV ranges between 40% and 100% and depends on the histological type.
Lymphoproliferations, referred to as post-transplant lymphoproliferative disorders, and classified histologically as diffuse large B-cell lymphoma, have been considered as EBV-driven lymphoproliferations occurring in the context of defective T-cell immunity against EBV.13
EBV is an established cause of extranodal NK/T-cell lymphoma (nasal type), previously called angiocentric T-cell lymphoma.2 Several recent large case series confirm the presence of EBV in tumour cells in nearly 100% of cases. Nasal NK/T-cell lymphoma cells exhibit several unique features including an absence of T-cell antigens and the expression of the NK cell marker CD56.11
Other T-cell lymphoproliferative disorders that have been reported to be associated with EBV include a subset of peripheral T-cell lymphomas, enteropathy-type T-cell lymphomas, γδ T-cell lymphomas (hepatosplenic and non-hepatosplenic), T-cell lymphoproliferative disorders after chronic EBV infection, EBV-associated cutaneous T-cell lymphoproliferative disorders (especially in Asia) and aggressive NK-cell leukaemias/lymphomas.
Angioimmunoblastic T-cell lymphoma is a distinct entity of peripheral T-cell lymphoma, which is also nearly always associated with EBV. Angioimmunoblastic T-cell lymphoma is characterised by prominent proliferation of high endothelial venules and follicular dendritic cells.11
Not all subtypes of classic HL (cHL) harbour EBV to the same degree. There are also data that suggest that the incidence of EBV-positive cHL is age-related.
Based on patterns of expression of the EBV genome, three types of latent gene expression have been reported: latency I, II and III.13 Whole gene expression and associated malignancies are reported in table 2.
Epstein-Barr virus latency pattern and associated malignancies
In AIDS-associated diffuse large B-cell lymphoma, viral gene expression patterns are variable, but the transforming EBV LMP1 is frequently expressed.13 ,14
In BL, EBV adopts the ‘latency I’ pattern, which involves only the expression of EBNA-1 and EBERs. The absence of EBV from the majority of sporadic or AIDS-associated BL indicates that it is not essential in the pathogenesis of this tumour. Conversely, the key pathogenic event in BL is the Ig/c-myc translocation found in all BL.3 ,15
In AIDS patients, HL is frequently associated with EBV infection (90–100% of cases) and can be considered as an EBV-driven lymphoma in which EBV-encoded LMP1 is thought to play a pivotal pathogenetic role. In these cases, in fact, the Hodgkin Reed-Sternberg cells, the neoplastic component of HL tumour, show consistent evidence of the EBV ‘latency II’ pattern, which involves expression of the transforming EBV protein LMP1, and EBNA-1 and LMP2A.12 ,16–19
In many PEL cases, the lymphoma cells harbour both EBV and KSHV, while some are only positive for KSHV.12 This event and the fact that the (non-transforming) EBV latency I pattern is expressed dually in EBV and KSHV positive PEL cells, suggests that KSHV is the driving force behind the development of PEL (figure 1).12
Kaposi's sarcoma-associated herpes virusKSHV belongs to the γ-herpesvirus family, and is the first known human member of the genus Rhadinovirus. This virus infects endothelial/spindle cells and lymphocytes.20 ,21 In KSHV-associated cancers, viral presence has been demonstrated by immunohistochemistry and by PCR in tumour tissues, but not in the adjacent tissues22 ,23 providing the initial evidences in favour of its involvement in the oncogenic process. The methods used to detect KSHV differ in sensitivity and in target detection, and detect only limited aspects of the oncogenic role of KSHV.24
Cancers that have been attributed to KSHV
A recent evaluation of the available epidemiological and molecular mechanistic evidence resulted in the classification of KSHV as a group 1 carcinogen. In the case of Kaposi's sarcoma (KS) and PEL the evidence supporting a causative role for KSHV was considered sufficient (table 1).
The epidemiological evidence supporting a causative role for KSHV in the pathogenesis of KS is strongly based on the fact that the virus is found in virtually all KS tumours, irrespective of the clinical form (‘classic’, African-endemic, HIV-associated, post-transplant)2 (figure 3).

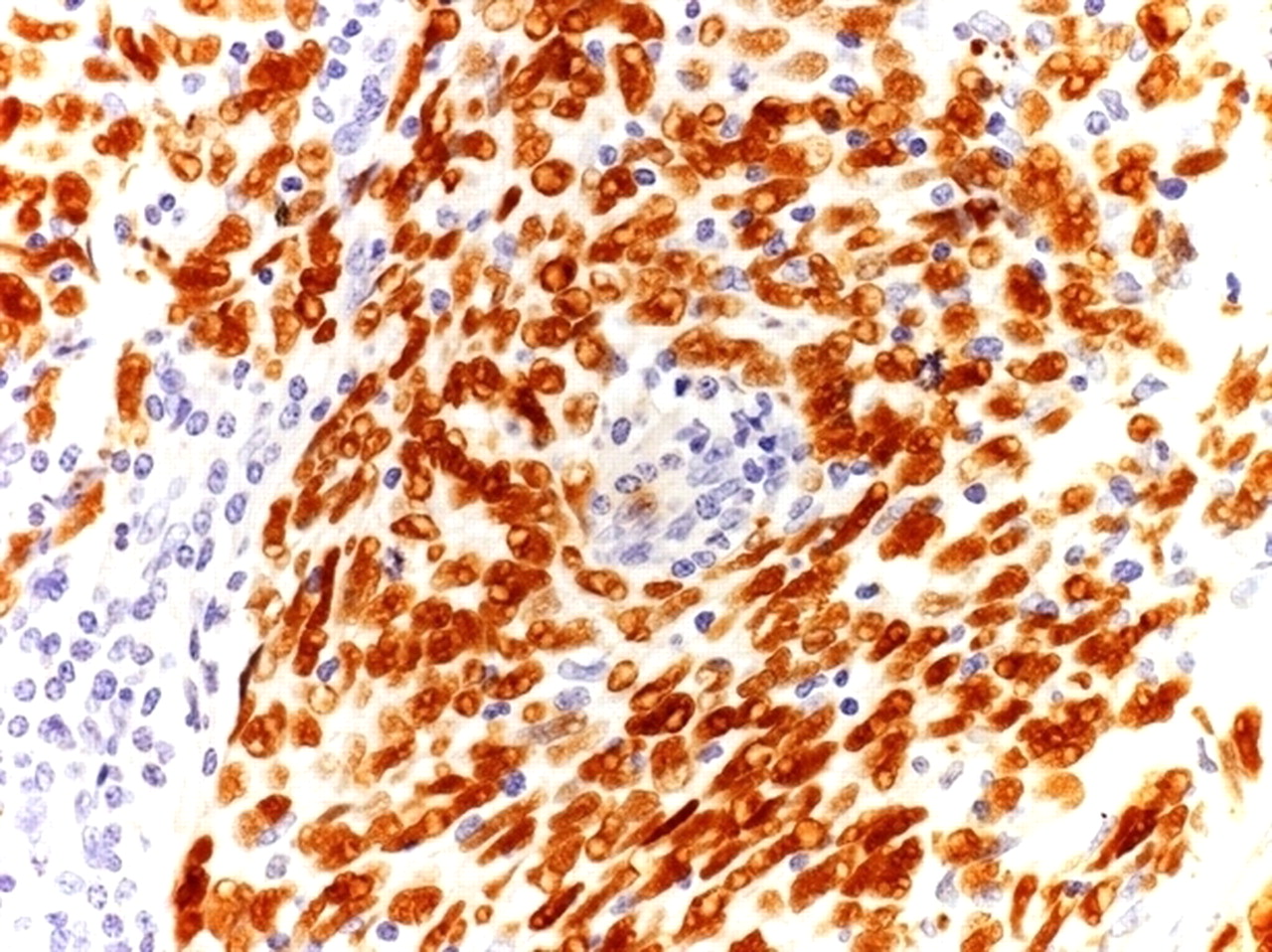
KSHV-associated Kaposi sarcoma (KS). Neoplastic spindle cells from a tumourous phase Kaposi's sarcoma display nuclear positivity KSHV with a speckled pattern of staining. KSHV positivity assessed by immunohistochemistry for ORF73/KSHV in formalin-fixed paraffin-embedded tissue section. Haematoxylin counterstain, original magnification ×40. KSHV, Kaposi's sarcoma associated herpes virus; ORF, open reading fragment.
PEL is encountered in AIDS patients and transplant recipients. Owing to its rarity, the epidemiological argument in favour of a causative involvement of KSHV is limited to the fact that KSHV is consistently found in this lymphoma. However, there are the following biological evidences: the same latent viral proteins (LANA, vcyc, vFLIP, kaposin) found in KS are also expressed in PEL; in addition, one of the KSHV interferon regulatory factor homologues, vIRF-3, shows a latent gene expression pattern in lymphoma cells and there is also substantial expression of an interleukin 6 homologue, vIL-6. Finally, experimental data show that the continuous expression of these viral proteins is required for PEL cell survival.11 ,25–27
Human papillomavirus
HPVs are small DNA viruses belonging to the family Papillomaviridae.28 ,29 HPV have circular, double-stranded DNA genomes typically containing 8 genes; the L1 gene encodes the principal capsidic protein, and its nucleotide sequence constitutes the basis for HPV classification into more than 120 ‘types’,28 while the L2protein is a minor virion component.30 ,31 The different HPV types are generally divided into those preferentially infecting the skin versus mucosal surfaces, the infection of the latter being associated with neoplastic diseases of the anogenital tract, and of head and neck squamous cell carcinomas.29 ,32 ,33 Among the genital HPVs, 15 types are considered ‘high-risk’ (in particular, HPV 16 and 18 that are responsible of >70% of the cancers), with over 99% of cervical lesions containing viral sequences.34
HPV infection requires that the virus passes through the epithelial basal layer and enters the basal cells, but the mode of entry is still debated.30–32 While many HPV types produce only productive infection, oncogenic HPV types contribute to the development of cancers, often arising at sites where productive infection cannot be supported.30
In these ‘high-risk’ HPV types, the E5, E6 and E7 genes are responsible for the production of the viral oncoproteins involved in the initiation and progression of cervical cancer.
Cancers that have been attributed to HPV
HPV infection-attributable cancers include carcinoma of the cervix, vulva, vagina, penis, anus, oral cavity and oropharynx and tonsil.11 The oncogenic types found most frequently in cervical cancer (HPV-16, 18, 31, 33, 35, 45, 52, 58) and four types less constantly found (HPV-39, 51, 56, 59) were classified in group 1 by the recent IARC evaluation.2 Of these, HPV-16 and 18 carry, by far, the highest risk of cancer.35–40 Several comprehensive studies have confirmed that these ‘high-risk’ HPV types cause virtually all cases of cervical cancer worldwide.39 ,40 HPV-16 is also the most important cause of anal cancer2 (figure 4).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
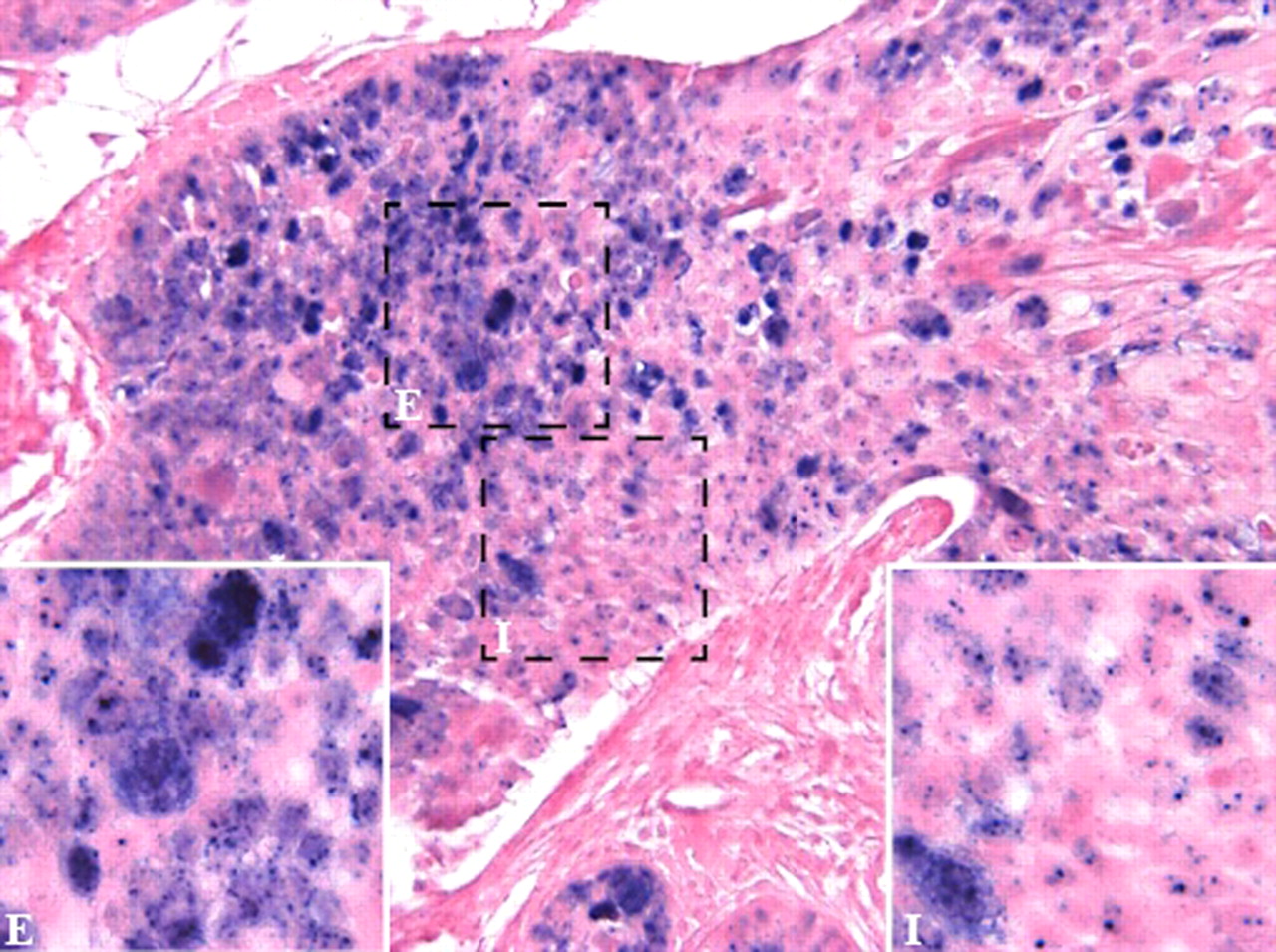
Human papillomavirus (HPV)-associated anal carcinoma. The figure shows an anal carcinoma in which neoplastic cells present ‘high-risk’ HPV infection either in episomal (E) or integrated (I) form. The positivity is nuclear. Inset: High magnification showing episomal (E) and integrated (I) pattern of staining. HPV positivity assessed by in situ hybridisation using a cocktail of probes recognising HR HPV types 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 68 and 70 in formalin-fixed paraffin-embedded section. Fast red counterstain, original magnification ×40, ×60 (inset).
Epidemiological data provide clues to the role of immune suppression in the development of HPV-related cancers on HIV setting.41 Yet, cervical cancer does not appear to have decreased after the introduction of highly active antiretroviral therapy.42 ,43
Pathological and virological techniques: clinical applications
EBV infection
The most abundant viral transcripts in latently infected cells, EBER1 and EBER2 (collectively called EBER), function to inhibit interferon-mediated antiviral effects and apoptosis. These two transcripts are expressed at around one million copies per latently infected cells. They are considered to be the best natural markers of latent infection. In situ hybridisation targeting one or both EBERs is the gold standard assay for reliably detecting EBV infection on formalin- and Bouin-fixed tumour samples.44 Today, this is a conventional technique to diagnose and classify NPC- and EBV-associated lymphomas in non-immunosuppressed hosts, and in AIDS-associated lymphomas and post-transplant lymphoid disorders.45
To identify the pattern of EBV latency expressed by tumour cells, the expression of EBV latent genes can be assessed by immunohistochemistry on fixed tissue samples by analysing EBV LMP1, LMP2a and EBV-encoded nuclear antigen 2.
The viral prevalence rates in tumours by using the various molecular methods are quite variable.46 Some studies employed PCR to amplify conserved regions of EBV DNA, but with this technique, false negative results may depend on the fact that PCR detects few genes at one time, while false positive results may be attributed to infiltrating lymphocytes, thus not allowing the distinction between lesion-specific EBV and background infection.47–50
Recent studies suggest that EBV genome chips covering the entire viral genome can optimally detect EBV infection in tumour cells46; EBV chips have the advantage of a low false positive rate and the simultaneous analysis of genome-wide viral gene expression pattern in tumour tissues. By using EBV chips, Yen et al 51 have demonstrated an increased sensitivity of EBV detection rate in oral cancer cells as compared with traditional molecular assays (from 15–77% to 82·5%).
Molecular methods have also demonstrated the monoclonal nature of EBV infection in EBV-related cancers.45
EBV DNA viraemia
Over the last few years, EBV DNA viral load testing has been extensively implemented for the prediction, diagnosis and monitoring of EBV-associated diseases.47 ,48 ,52 ,53 EBV DNA normally resides inside circulating B lymphocytes, whereas patients with EBV-associated cancers also have detectable viral DNA in plasma, making the detection of extracellular viral load a suitable specimen type for clinical purposes.48 ,53
In EBV-related cancers, viral load may be produced through different mechanisms and have peculiar significance in cancers of different origin. At least two different mechanisms may be hypothesised. In the first, including cancers arising in immunocompetent hosts (NPC, HL, NK/T cell lymphomas), the EBV-DNA detectable in the peripheral blood is not related to viral reactivation, but rather, derives directly from the necrosis/apoptosis of tumour cells.54 ,55 For this reason, it may be considered as a real tumour biomarker.
In areas where NPC is frequent, anti-viral capsidic antigen IgA antibody test is used to screen populations at higher risk to develop the tumour. Serological assays are cheap and technically easy, but suffer from limited specificity. For this reason, EBV DNA has been proposed as a confirmatory test, when immunoglobulin A-viral capsidic antigen results are positive or indeterminate.56 EBV DNA assay may be also used as a diagnostic tool, for example, when histology gives inconclusive results. In high-incidence areas, plasma EBV DNA has a diagnostic sensitivity of 95%,56 while in low-incidence regions, sensitivity is reduced to 70–75%.51 ,57 Some authors tried to support the usefulness of EBV DNA determination in nasopharyngeal brushings, rather than in the peripheral blood.58
Pre-therapy EBV DNA is an independent prognostic factor to international union against cancer staging in NPC.59 In fact, several studies have demonstrated that overall survival, or relapse-free survival, were statistically significantly lower in NPC patients with elevated EBV DNA.59 ,60 In order to identify prognostic factors related to therapy, Lo et al demonstrated that EBV DNA declined after radiotherapy49; however, plasma EBV DNA levels depend on the balance between production and catabolism, and limited information is available at the moment on these issues. In agreement with histopathological data, EBV DNA is detectable in all EBV positive HL before therapy,61 ,62 and in one study predicts response to therapy,62 but the use of EBV DNA as a diagnostic or predictive marker in HL patients is not recommended.
In the second type of cancers, including EBV-related lymphoproliferative disorders, EBV DNA is actively released from circulating EBV-positive B cells proliferating because of the absence of an effective T-cell control.63 In this case, EBV DNA level may not be considered as a true tumour marker, but rather a measure of immunosuppression. Having in mind this limitation, EBV viral load is considered as very useful to predict, diagnose and monitor post-transplant lymphoproliferative disorders.48
Approximately 20% of HIV positive asymptomatic patients have a detectable viral load in the peripheral blood, and it has been hypothesised that EBV DNA may be used as a reliable predictor of the development of non-HLs, but detailed longitudinal studies failed to support this hypothesis.64 ,65
The methods currently used to measure EBV DNA consistently differ for the EBV genome regions detected, for the detection techniques used and for the type of external standards used to set up the standard curve, producing a significant variability among laboratories.47 ,53 Despite these limitations, EBV viral load is considered relevant for the management of EBV-associated cancers.
KSHV infection
The detection of KSHV infection in tumour cells is the prerequisite for a precise diagnosis of KS or PEL. KSHV infection can be assessed on formalin-fixed cytospin (or cell blocks) obtained from effusions or formalin-fixed tissue sections.12 ,66
The presence of KSHV can be ascertained by immunohistochemistry on cells or sections using antibodies against the KSHV latency-associated nuclear antigen encoded by viral open reading fragment (ORF) 73. As KSHV in tumour cells is in latent phase, ORF73 immunohistochemistry is the gold standard to reliably detect KSHV infection in tumour samples.12 This antigen is detectable in the nucleus of almost all the infected cells. Positive cases can also be tested for viral interleukin-6 (vIL-6), a KSHV cytokine homologue. Staining for vIL-6 is cytoplasmic and is restricted to a limited number of infected cells. Positive controls for ORF73 consist of KS biopsy samples, whereas positive controls for vIL-6 consist of AIDS-associated multicentric Castleman's disease (MCD) biopsy samples.12
The presence of KSHV can be confirmed by PCR analysis of three KSHV regions (ie, K330233, ORFK9-3, ORF72) using primer sequences.66
KSHV viraemia
During KSHV infection, viral DNA can be detected both in peripheral blood mononuclear cells and as cell-free DNA (plasma viraemia)67 and may be used to predict, diagnose or prognosticate diseases associated with this virus.
Engels et al have demonstrated that, among individuals with HIV-KS co-infection, KSHV viraemia predicts the further development of KS,68 ,69 while no conclusive studies are presently available on the predictive role of viraemia in the development of KS in the HIV-negative population.70
There is a wide consensus in the scientific literature that KSHV viraemia does not have a diagnostic role in any of the KSHV-associated diseases,71 while several studies have considered KSHV viraemia as a useful prognostic indicator of disease progression and/or shorter survival in KS70 ,72 and PEL.73
Finally, KSHV viraemia has been demonstrated to be a reliable method to monitor the response of KS to highly active antiretroviral therapy 67 ,74 or to anti-tumour chemotherapy in MCD.75 ,76
HPV infection
In developed countries, a cervical Papanicolaou (Pap) test (a screening test to detect cervical cell changes) is still used to detect abnormal cells that may develop into cancer. On the other hand, genital warts that are the only visible sign of ‘low-risk’ HPV continue to be identified with a visual check.
An HPV test for cervical infection as a follow-up for women who have an ambiguous Pap test for general cervical cancer screening has recently been adopted. This HPV test (HC2) can identify ‘low’ and ‘high-risk’ types of HPV associated with the development of cervical cancer. However, today it is possible to test for HPV DNA in extracervical infections by means of multiple approaches. Many assays for HPV detection are available today, each characterised by its own analytical sensitivity. In situ hybridisation can also be applied to detect HPV in slides (scrapes or tissue sections). There are two commercially available systems able to detect HPV in tissue samples, that is, the Ventana System (Ventana Systems, Inc., Harvard, MA, USA), able to identify ‘high-risk’ HPV types, whereas Dako Probe (Dako, Denmark) is specific for detecting HPV16. Both methods allow the identification of the integrated and episomal form. P16 immunohistochemistry has been advocated as a surrogate marker of ‘high-risk’ HPV infection. Finally, there are the PCR-based assay systems, often linked to a specific genotyping system.
With the presently available assays that have extremely high analytical sensitivities, there is the risk that the test is too sensitive and detects virus without underlying biological or clinical consequences. There is sufficient evidence that increased analytical sensitivities of assays result in decreased clinical specificities (ie, more positive test results in the absence of virus-induced disease). Thus, a positive ‘high-risk’ HPV test does not automatically mean that the virus has been involved in the initiation or maintenance of carcinogenesis.77
HPV DNA may be present in cervical lesions in the absence of any overt disease, and may represent a silent viral infection, possibly because of an efficient immune control at the mucosal surface.
Since HPV DNA integration is found in most invasive cancers, it has been considered as a step promoting viral carcinogenesis; however, experimental data are conflicting, and this topic is still debated.78 ,79 The quantitative measurement of HPV DNA integration in human cells may offer information to classify HPV-related cancers based on molecular characteristics, but the significance of quantitative data is still debated, since viral load values are an average summed over many infected and uninfected cells.78
Owing to the fact that viral integration disrupts the E2 gene, while the E6 gene is maintained, the E2/E6 ratio accurately measures the amount of integrated HPV in human tumour samples.80 HPV E6/E7 mRNA testing may also be a potentially useful biomarker to evaluate the risk of progression of HPV-related pre-neoplastic lesions.81
Since HPV-associated cancers are associated with ‘high-risk’ viral types, HPV typing is essential for diagnostic and research purposes. Typing can be performed by amplifying HPV L1 gene combined with reverse line blotting assay of sequencing.78
Several studies suggest that in a proportion of HPV-related lesions, several viral types are simultaneously detected78 ,81 However, there is a lack of consensus in literature about the prevalence and carcinogenic role of co-infections with multiple HPV types in cervical and head and neck carcinomas.82
In conclusion, virological assays to detect HPV may measure the qualitative and quantitative presence of HPV DNA and determine the HPV-infecting type.
In the future, molecular assays on laser-microdissected single tumour cells may be essential to study the viral oncogenic pathways and to obtain more reliable diagnostic and prognostic information on HPV-related cancers.
Future scenario
Owing to the fact that a fraction of malignancies can be caused by chronic infections, vaccination may be effective against the onset of these cancers. The future scenario we may have to face ahead of us in case the development of vaccines were feasible, and vaccination campaigns were effective, may consist of a 15–20% reduction of human malignant tumours worldwide.
In conclusion, the future scenario is promising. Yet, several issues relating to viral agents and their role on carcinogenesis are still open, and tumour prevention and treatment. There is still much to do in these fields as far as research is concerned. First, we need to investigate whether some neoplastic diseases with limited evidence (table 3) are only coincidentally bound to such biological agents. We are referring to carcinoma of the larynx and HPV, carcinoma of the stomach and EBV, MCD and KSHV. Moreover, other biological agents are still under investigation to establish whether they belong to group 1, that is, to cancer-causative biological agents. Therefore, the fight against cancer both in developing and developed countries can benefit from this knowledge, specifically of the induction of complete protection against primary infection, or prevention of chronic infections and of the disease. It is still doubtful whether which of the two is the most effective/feasible way.
Biological agents assessed by the International Agency for Research on Cancer monograph working group (adapted from Bouvard et al 2)
Interactive multiple choice questions
-
This JCP review article has an accompanying set of multiple choice questions (MCQs). To access the questions, click on BMJ Learning: Take this module on BMJ Learning from the content box at the top right and bottom left of the online article. For more information please go to: http://jcp.bmj.com/education. Please note: The MCQs are hosted on BMJ Learning—the best available learning website for medical professionals from the BMJ Group. If prompted, subscribers must sign into JCP with their journal's username and password. All users must also complete a one-time registration on BMJ Learning and subsequently log in (with a BMJ Learning username and password) on every visit.
Take home messages
-
Infectious agents are implicated as causative factors in a significant proportion of human cancers. Due to their direct carcinogenic role, EBV, KSHV, and HPV are particularly important among viruses. Therefore the use of well-established pathological and virologic techniques is helpful to diagnose and characterize viral-associated cancers. However, several issues relating to viral related cancers are still open including tumour prevention and treatment.
References
Footnotes
-
Search strategy and selection criteria References for this review were identified through searches of PubMed for articles published from January 1992 to August 2010, by use of the terms vaccination, viral agents, carcinogenesis, oncogenic viruses. Articles from IARC Monographs have also been reviewed.
-
AC has been a member of WHO IARC Monograph Working Group on Biological Agents, Lyon—2009.
-
Competing interests None.
-
Provenance and peer review Not commissioned; externally peer reviewed.