Article Text

Abstract
Background/Aim: It has been reported previously in cases of adenosquamous carcinoma of the lung in Okinawa, a subtropical island 2000 km south of mainland Japan, that the squamous cell carcinoma components were positive for human papillomavirus (HPV) by non-isotopic in situ hybridisation (NISH). The adenocarcinoma cells adjacent to the squamous cell carcinoma components were enlarged and also positive for HPV. This is thought to indicate that after adenocarcinoma cells are infected with HPV, they undergo morphological changes, and that “squamous metaplasia” follows. In this present study, the effects of HPV transfection into adenocarcinoma cells were examined. The relation between the region expressing the HPV gene and squamous metaplasia was also studied.
Methods: Plasmid pBR322 containing HPV type 16 (HPV-16) was transfected into cultured colonic adenocarcinoma (DLD-1) and lung adenocarcinoma (PC-14) cells using the calcium phosphate method. Neomycin was used as a selection marker. The presence of HPV E1, E2, E4, E5, E6, E7, L1, and L2 mRNAs and also transglutaminase 1, involucrin, cyclin dependent kinases (CDKs), cyclins, caspases, apoptosis inducing factor, DNase γ, Fas, and Fas ligand mRNAs in HPV transfected cells was investigated by means of reverse transcription polymerase chain reaction (RT-PCR). The G0–G1 cell population was analysed by flow cytometry. Morphological examination under light and electron microscopes was also carried out.
Results: The virus transfected cells showed squamous metaplasia when they were injected into severe combined immunodeficient mice, expressing the high molecular weight keratin (Moll’s number 1 keratin) and involucrin molecules immunohistochemically, and involucrin and transglutaminase I mRNAs by RT-PCR. The squamous metaplasia was most conspicuous in the HPV transfected DLD-1 cell when compared with HPV transfected PC-14 cells. Squamous metaplasia was most clearly demonstrated in one HPV transfected DLD-1 cell clone, which expressed not only E2 but also E6–E7 fusion gene mRNA. Viral L1 mRNA expression was absent in HPV transfected cell clones, and was not related to squamous metaplasia. The growth rate of HPV transfected cells was reduced. Transfection of the virus into the cultured adenocarcinoma cells increased the G0–G1 cell population greatly, as assessed by flow cytometer analysis. Furthermore, in the virus transfected cells, apoptosis was also observed by means of the terminal deoxynucleotidyl transferase mediated dUTP biotin nick end labelling method.
Conclusion: HPV transfection into adenocarcinoma cells induced clear squamous metaplasia. One of the HPV transfected cell clones that expressed E2 and E6–E7 fusion gene mRNA showed the squamous metaplasia particularly clearly, and apoptosis was also demonstrated.
- human papillomavirus
- gene transfection
- squamous metaplasia
- apoptosis
- BPV, bovine papillomavirus
- CDK, cyclin dependent kinase
- HMK, high molecular weight keratin
- HPV, human papillomavirus
- NISH, non-isotopic in situ hybridisation
- PAGE, polyacrylamide gel electrophoresis
- PAS, periodic acid Schiff
- PBS, phosphate buffered saline
- PCNA, proliferation cell nuclear antigen
- PMSF, phenylmethylsulphonyl fluoride
- RT-PCR, reverse transcription polymerase chain reaction
- SCID, severe combined immunodeficient
- SDS, sodium dodecyl sulfate
- TUNEL, terminal deoxynucleotidyl transferase mediated dUTP biotin nick end labelling
Statistics from Altmetric.com
- BPV, bovine papillomavirus
- CDK, cyclin dependent kinase
- HMK, high molecular weight keratin
- HPV, human papillomavirus
- NISH, non-isotopic in situ hybridisation
- PAGE, polyacrylamide gel electrophoresis
- PAS, periodic acid Schiff
- PBS, phosphate buffered saline
- PCNA, proliferation cell nuclear antigen
- PMSF, phenylmethylsulphonyl fluoride
- RT-PCR, reverse transcription polymerase chain reaction
- SCID, severe combined immunodeficient
- SDS, sodium dodecyl sulfate
- TUNEL, terminal deoxynucleotidyl transferase mediated dUTP biotin nick end labelling
Metaplasia is the process whereby one type of differentiated cell is replaced by another, usually in response to abnormal stimuli.1 “Squamous metaplasia” of the bronchial mucosa and “intestinal metaplasia” of the gastric mucosa are well known and commonly accepted phenomena. As reported previously2,3 in adenosquamous carcinoma cases of the lung, the squamous cell carcinoma components are positive for human papillomavirus (HPV) by non-isotopic in situ hybridisation (NISH). The adenocarcinoma cells adjacent to the squamous cell carcinoma components are enlarged, and also show positivity for HPV by NISH. The production of high molecular weight keratin (HMK) and involucrin molecules (markers of the cornified squamous cell) is induced in the squamous cell carcinoma components and also in the adjacent enlarged cells.3–12 This is thought to indicate the transition from adenocarcinoma to squamous cell carcinoma. We postulated3 that after adenocarcinoma cells are infected with HPV, they undergo morphological changes, expressing HMK and involucrin, and that squamous metaplasia follows. Metaplasia is the change in the differentiation pathway to which the stem cell progeny commit. Squamous metaplasia is the common adaptive response to chronic irritations in non-neoplastic tissue. A similar process of squamous metaplasia is common in neoplasms, particularly in adenocarcinomas of the lung, uterus, etc. Metaplasia is said to occur in tumours, but the term metaplasia is better used for changes in non-tumorous tissues. The squamous differentiation is generally used when a squamous cell carcinoma component is seen in the adenocarcinoma.13 However, in our report squamous metaplasia is used to indicate the transition from adenocarcinoma cell to squamous carcinoma cell morphology, including the expression of the appropriate molecular markers—for example, cytokeratins, involucrin, and transglutaminase I.
Tinsley et al reported HPV type 1 transgenic mice in which the E1–E4 protein was detected in the upper suprabasal layers of the skin in paws and tail.14 A 1.7 kb RNA sequence corresponding to the E6 and E7 transcript was prominent in tails. In such transgenic mice the epidermis of the tail showed hyperplasia, with both hyperkeratosis and focal parakeratosis. Moreover, based on histological examination of certain human skin lesions, such as verruca vulgaris and condyloma acuminatum, HPV has been found to cause keratinisation of the skin. It is considered that a region of the viral genome causes cell differentiation.
“In adenosquamous carcinoma cases of the lung, the squamous cell carcinoma components are positive for human papillomavirus by non-isotopic in situ hybridisation”
In our study, transfection of HPV-16 into cultured adenocarcinoma cells was carried out. The choice of HPV-16 reflects the previous finding2 that squamous metaplasia was more apparent in the case of HPV-16 transfected cells than in HPV-6 and HPV-18 transfected cells. Furthermore, the relation between regions expressing HPV gene mRNA and squamous metaplasia is examined, as is the possibility that HPV plays an important role in squamous metaplasia.
MATERIALS AND METHODS
Cells
The cultured human intestinal adenocarcinoma (moderately differentiated) cell line, DLD-1, and human lung adenocarcinoma (poorly differentiated) cell line, PC-14, were obtained from the Cancer Research Resources Bank, National Institute of Health, Japan (Setagaya, Tokyo, Japan). DLD-1 cells are adherent, and PC-14 cells were weakly adherent to culture dishes. Both were cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum (Bio Whittaker, Walkersville, Maryland, USA) at 37°C in a humidified atmosphere of 5% CO2/95% air. The characteristics of both cells have been reported elsewhere.5,15 We tested further characteristics of the cells. Overexpression of Bax and Bcl-2 was not demonstrated immunohistochemically. Sequence analysis of the p53 gene in DLD-1 revealed the mutation at Ser241Phe (C936T), as reported previously16 (data not shown), but no mutation was demonstrated in PC-14 cells. CaSki cells,17 which are HPV-16 infected epidermoid orvical carcinoma cells, were used as a control.
HPV DNA
HPV-16 in plasmid pBR322 was obtained from the Japanese Cancer Research Resources Bank (National Institute of Health, Division of Genetic Resources, Shinjuku, Tokyo, Japan) with the permission of Dr Zur Hausen. Neomycin was used as a selection marker. The neomycin resistant gene in pMC1neo poly A+ was obtained from Stratagene (La Jolla, California, USA).
The reconstructed plasmid pBR322, which has HPV-16 at the Hind III site and the neomycin resistance gene at the BamHI site, was prepared as reported previously.2
Transfection of HPV DNA into cultured cells
Using Chen and Okayama’s calcium phosphate method,18 HPV DNA was transfected into DLD-1 and PC-14 cells and 5 × 105 cells were seeded in a 10 cm culture dish containing 10 ml of fresh growth medium and incubated overnight at 37°C in 3% CO2. The HPV DNA–CaCl2 was added dropwise to the dish containing the growing cells, swirled to mix well, and incubated for 10 hours at 37°C in 5% CO2.
HPV DNA–CaCl2 was made as follows: HPV DNA (1 mg/ml in TE buffer (10 mM Tris/HCl, pH 7.5, containing 1mM EDTA)) was added to 0.25M CaCl2 in sterile distilled water. Then, 0.5 ml 2× HBS buffer (280mM NaCl, 1.5mM Na2HPO4, 50mM HEPES, pH 7.1) was added dropwise to the mixture with constant swirling, and left at room temperature for 20 minutes.
After treatment with 15% glycerol, HPV transfected cells were re-fed with fresh growth medium and incubated at 37°C for 24 hours in 5% CO2. Thereafter, the cells were submitted to a selection process in 200 μg/ml of G418 (Geneticin; Gibco BRL, Grand Island, New York, USA). Four weeks later, a single cell was subcultured from the surviving colonies using 96 well multiplates.
Thirteen clones of HPV transfected DLD-1 cells and 12 clones of HPV DNA transfected PC-14 cells were obtained. These HPV transfected cells were cultured. After 10 passages, their characteristics were examined.
Growth curve and cell cycle analysis
A 0.2 ml aliquot of 106/ml HPV transfected cells was seeded into 35 mm culture dishes. After 1, 2, 3, 5, 7, 10, 14, and 20 days the cells were counted using a haemocytometer after trypan blue staining, and the growth curve was obtained.
For the cell cycle analysis, the DNA content was measured by means of flow cytometry (Becton Dickinson, Mountain View, California, USA). HPV transfected cells were harvested and fixed in cold 70% ethanol for 30 minutes on ice. After washing in phosphate buffered saline (PBS), pH 7.0, cells were pelleted and resuspended in 1 ml of 50 μg/ml propidium iodine in PBS containing 20 μg/ml of RNase and 0.05% Triton X-100 for a further 30 minutes, and then diluted in PBS containing 1% bovine serum albumin and immediately analysed by flow cytometer.
Injection of HPV transfected cells into SCID mice
HPV transfected DLD-1 cells and PC-14 cells (107 cells) were injected subcutaneously into the backs of severe combined immunodeficient (SCID) mice19 (CB17/Icr-Scid Jcl), which were obtained from Japan Bio Co (SEAC Yoshitomi Co, Fukuoka, Japan) four weeks after birth. All animals used in our study were maintained and conditioned in the Institute for Animal Experiments, Ryukyu University School of Medicine for one week before the experiments, and all experimental protocols were approved by the Animal Care and Use Committee, Ryukyu University. The SCID mice into which HPV transfected DLD-1 and PC-14 cells were injected were previously injected intraperitoneally with 0.3 ml of 30 times diluted anti-asialo GM1 ganglioside antibody19 (Wako, Tokyo, Japan) three times/day.
Morphological examination
HPV transfected cultured cells on the dishes were examined under a phase contrast microscope. HPV transfected cultured cells and tumours, which were obtained four weeks after injection of transfected cells into mice, were fixed in 10% sodium phosphate buffered formalin, pH 7.2. After fixation, HPV transfected cells and tumours were subjected to routine histological examination. Haematoxylin and eosin, periodic acid Schiff (PAS), and alcian blue staining were performed on the cultured cells on the dishes and 4 μm dewaxed sections of the tumour cells. The antibody to involucrin (a marker for keratinocyte differentiation) was obtained from Sigma (St Louis, Missouri, USA), and antibody to high molecular keratin (HMK) (Moll’s10 No. 1, 5, 10, 14), proliferation cell nuclear antigen (PCNA), Ki-67, and p53 protein from Dako (Carpinteria, California, USA). Anticaspase 4 antibody was from Wako (Osaka, Japan). Immunohistochemical studies were carried out by the avidin–biotin–peroxidase method. Apoptosis was detected using the TUNEL (terminal deoxynucleotidyl transferase mediated dUTP biotin nick end labelling) method. The TUNEL method was performed with a Takara in situ apoptosis detection kit (Takara, Otsu, Japan), according to the manufacturer’s instructions.
NISH was performed on all specimens using HPV-16 biotin labelled probes from the ENZO PathoGene in situ HPV tissue hybridisation kit (Farmingdale, New York, USA). NISH was carried out according to Cooper and colleagues20 and the manufacturer’s instructions. Biotin was detected using alkaline phosphatase labelled streptavidin. 5-Bromo-4-chloro-3-indolyl phosphate and nitrobluetetrazolium were used as substrate and chromogen, respectively. For the immunohistochemical studies, TUNEL method, and NISH, non-HPV transfected DLD-1 and PC-14 cells were used as controls.
For electron microscopic examinations, samples were fixed in buffered 2.5% glutaraldehyde and postfixed in 1% osmium tetroxide, then embedded in Epon 812 resin in the usual manner. Ultrathin sections were observed using the Hitachi H7500 electron microscope (Hitachi Ltd, Tokyo, Japan).
Purification of involucrin and western blot analysis
Involucrin molecules were isolated according to the method of Schmiedeknecht and colleagues21 and Yaffe et al.12 HPV transfected cultured cells were washed twice with PBS, pH 7.2, containing 20mM EDTA, once with PBS containing 20mM EDTA and 0.2 mM phenylmethylsulphonyl fluoride (PMSF), and were then harvested by scraping. The cells were collected in PBS containing 20mM EDTA and pelleted by centrifugation; they were then disrupted by two freeze–thaw cycles in 10 volumes of 10mM Tris/HCl, pH 7.4, 10mM EDTA, containing 0.3 mg/ml PMSF, 5 μg/ml leupeptin, and 0.1 μl/ml pepstatin. The sample was centrifuged at 10 000 ×g for 10 minutes at 4°C. The supernatant was collected and 1/10 volume of 100% trichloroacetic acid (TCA) was added. Thereafter, the pellet was obtained by centrifugation at 27 000 ×g for 20 minutes at 4°C, and dissolved in 9M urea, 2% Triton X-100, and 5% 2-mercaptoethanol. After sonication (one minute, three times), a 1/4 volume of 10% sodium dodecyl sulfate (SDS) was added. The samples were electrophoresed on an 8.5% acrylamide gel, transferred to a nitrocellulose membrane, and incubated with anti-involucrin antibody. Then they were visualised by incubating with H2O2 and 3,3′ diaminobenzidine, after incubation with a second antibody (antimouse rabbit immunoglobulin; Dako, Kyoto, Japan) labelled with peroxidase.
In the case of HPV transfected cell tumours in the SCID mice, the samples were homogenised using a Polytron homogeniser (Kinematica GmbH, Steinhofhalde, Switzerland) in PBS containing 20mM EDTA and 0.2mM PMSF, and then centrifuged for five minutes at 15 000 ×g. The samples were disrupted by two freeze–thaw cycles, and were processed by the same procedures as described above.
Purification of the keratin molecules and western blot analysis
The keratin molecules were extracted according to the method of Franke and colleagues22 and Katagata et al.9 The HPV transfected culture cells were washed twice with PBS and scraped with a rubber policeman. The tumours obtained from SCID mice were minced. Five to 10 volumes of 10mM Tris/HCl, pH 7.4, containing 10mM EDTA, 0.3 mg/ml PMSF, 5 μg/ml of leupeptin, and 0.1 μl/ml of pepstatin were added to the samples, which were then sonicated three times for four minutes each, followed by centrifugation for 10 minutes at 10 000 ×g. After repeating the above procedure three times, the pellets were dissolved with equal volumes of 9M urea, 10% 2-mercaptoethanol, and 4% NP40, at 37°C for six hours. Thereafter, the samples were analysed using two dimensional polyacrylamide gel electrophoresis (PAGE). After SDS-PAGE was carried out according to the method of Laemmli,23 two dimensional PAGE was performed as described by O’Farrel,24 using Pharmalyte (Amersham Pharmacia Biotech AB, Uppsala, Sweden), pH 3–10. After electrophoresis, the samples were transferred on to nitrocellulose membranes and the membranes were stained with anti-HMK (Moll’s10 numbers 1, 5, 10, 14) antibody (Dako, Carpinteria, California, USA).
Detection of HPV mRNA by RT-PCR
Total RNA was extracted from the samples according to the standard acid/guanidium/phenol/chloroform method. A sample (the cells in 10 cm dish) was homogenised with 500 μl of solution (4M guanidium thiocyanate, 25mM sodium citrate, 0.5% N-lauroylsarcosinate, and 0.1% 2-mercaptoethanol, pH 7.0), then 50 μl of 2M sodium acetate, 500 μl of water saturated phenol, and 100 μl of chloroform/isoamylalcohol (24/1 (vol/vol)), pH 4.5, was added. The sample was cooled on ice for 15 minutes, and centrifuged at 10 000 ×g at 4°C for 20 minutes. A 600 μl aliquot of ice cold isopropanol was added to the aqueous phase, which was then kept in a −20°C freezer for two hours. The RNA was obtained by centrifugation at 10 000 ×g for 30 minutes. The sample was digested by means of DNase (Takara). The glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene was amplified by primers designed around introns to produce a fragment of 2086 bp from DNA and 234 bp from RNA.6 The sense primer AGGTGAAGGTCGGAGTCAACG (nucleotide position, 1460–1480) and antisense primer GCTCCTGGAAGATGGTGATGG (nucleotide position, 3542–3412) and Takara Ex Taq DNA polymerase (Takara) were used for the PCR. The genomic 2086 bp GAPDH DNA was not amplified. Then the RNA was reverse transcribed at 42°C for 60 minutes in a 20 μl reaction volume using a First Strand cDNA synthesis kit (Clontech Lab, Palo Alto, California, USA), according to the manufacturer’s instructions. cDNA was incubated at 95°C for five minutes to inactivate the reverse transcriptase, and used as template DNA in the PCR amplification of the HPV-16 E1, E2, E4, E5, E6, E7, L1, and L2 regions. The primers and probes for these HPV E1, E2, E4, E5, E6, E7, and L1 or L2 regions are shown in table 1 and fig 1. The PCR conditions were the same as reported previously.3,26
Primers and probes used for detection of human papilloavirus type 16 (HPV-16) DNA by the polymerase chain reaction

Positions of the primers used for the detection of human papillomavirus type 16 DNAs by the polymerase chain reaction.
Detection of other mRNAs of interest by RT-PCR
Using cDNA reverse transcribed from RNA as described above, transglutaminase I,27 involucrin, Fas, Fas ligand, bax, bcl-2,28 p16α, and p16β,29 caspase 3,30 caspase 8,31 caspase 9,30 AIF,32–34 DNase γ,35 DNA fragmentation factor 45, cyclin D1, D2, and D3, cyclin A,36 cyclin E,37 and cyclin dependent kinases (CDKs) 2, 4, and 636 were detected by the RT-PCR method. The RT-PCR primers for tranglutaminase I, Fas, Fas ligand, bax, bcl-2, p16α and p16β, caspase 3, 8, and 9, DNase γ, cyclin D1, D2, and D3, cyclin A and E, CDK2, 4, and 6 were the same as in previous reports. Table 2 lists the primers and probes used. The PCR conditions were similar to those described earlier.27–30,35–37 The template cDNA concentrations of the samples were adjusted according to the RT-PCR product concentration of GAPDH.
Primers used for characterisation of the human papillomavirus transfected cells
Negative controls for PCR analysis
To prevent false positive results, the sample preparation and the PCR were carried out by two authors independently (TK and KK). Furthermore, distilled water was used as a negative control and no positive reaction was obtained.
Sequence analysis of PCR products
PCR products were extracted from the agarose gel. The extracted DNA was cloned into T-vector prepared from pBluescript II (Stratagene), according to the method of Marchuk et al.38. The pBluescript plasmid was digested with EcoRV, and incubated with Taq (Cetus-Takara) (1 U/μg plasmid) using standard buffer conditions: 10mM Tris/HCl, pH 8.3, 50mM KCl, 1.5mM MgCl2, and 200 μg/ml bovine serum albumin in the presence of 2mM dTTP for two hours at 70°C. Sequence analysis was carried out using the Hitachi SQ 5500 DNA sequencer (Hitachi).
Restriction enzyme cleavage and blot hybridisation
Between 10 and 20 μg of DNA were digested with the appropriate restriction endonuclease, and after electrophoresis on a 0.8% agarose gel, the DNA was denatured and transferred to a nitrocellulose filter. Filters were hybridised with HPV DNA.39
RESULTS
Growth curve and cell cycle analysis
Thirty HPV-16 transfected DLD-1 cell clones and 12 HPV-16 transfected PC-14 cell clones were obtained. All types of HPV transfected DLD-1 cells adhered to the culture dishes, as they had before HPV transfection. In contrast, PC-14 cells, which were only weakly adherent before the viral transfection, became tightly adherent to the dishes.
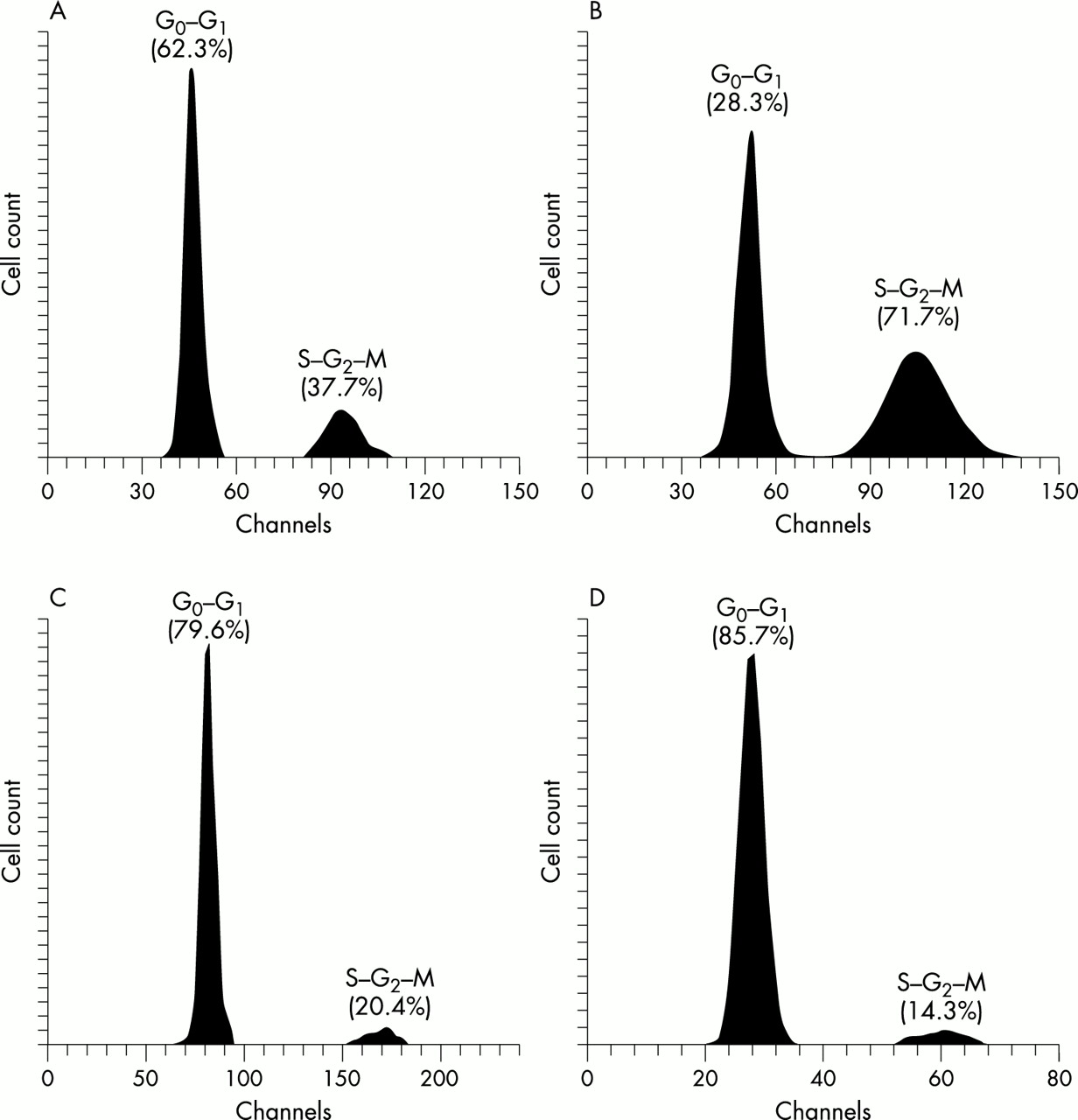
The growth rate of HPV transfected cells was reduced. Before HPV transfection, the G0–G1 phase cell populations in the DLD-1 and PC-14 cells were less than 62.3% and 28.3%, respectively (fig 2A,B), and these cell populations increased after transfection. Among both the DLD-1 and PC-14 HPV transfected cell clones, the G0–G1 cell populations were particularly high in the 16Da clones, which expressed E2 and E6–E7 fusion gene mRNA, and 16Pa clones, which expressed E6 and E7 gene mRNA, reaching about 80% (79.6% and 85.7%, respectively) (fig 2C,D). However, cells expressing only one of E2, E6, or E7 had slightly lower G0–G1 cell populations of about 70%.

Flow cytometric analysis. (A) G0–G1 cell population of cultured DLD-1 cells without human papillomavirus (HPV) DNA transfection (control); 62.3% were G0–G1 phase cells; 37.7% were S–G2–M phase cells. (B) G0–G1 cell population of cultured PC-14 cells without HPV DNA transfection (control); only 28.3% of cells were G0–G1 phase; 71.7% were S–G2–M phase cells. (C) G0–G1 cell population of HPV-16 transfected DLD-1 cells (16Da cells); 79.6% of cells were G0–G1 phase cells; 20.4% were S–G2–M phase cells. (D) G0–G1 cell population of HPV-16 transfected PC-14 cells (16Pa cells); 85.7% of cells were G0–G1 phase; 14.3% were S–G2–M phase cells.
Detection of HPV E1, E2, E4, E5, E6, L1, and L2 mRNA, in addition to mRNA of other molecules of interest
The HPV-16 transfected 16Da clone (one of the DLD-1 cell clones), which showed the clearest squamous metaplasia, in addition to all of the HPV-16 transfected DLD-1 and PC-14 cell clones that also showed squamous metaplasia were analysed for mRNA (table 3). In the present HPV-16 transfected cells (16Da, 16Db, and 16Pa), the previously reported splicing sites21,40–42 at 226, 409, and 526 were not demonstrated. In the case of 16Da cells, using the primer pair 16E6-a and 16E7-b, the expected 695 bp product was amplified (fig 3). However, the primer pair 16E7-a and 16E2-b produced no corresponding PCR product. Using the primer pairs 16E6-a/16E7-b and 16E4-a/16E5-b the expected PCR products were amplified (fig 3). The L1 region was not amplified in 16Da. One HPV transfected cell, 16Db, which expressed only E4 and E6 mRNA, and not L1 mRNA, was used for further investigations.
Detection of mRNAs in human papillomavirus type 16 (HPV-16) transfected DLD-1 and PC-14 cells

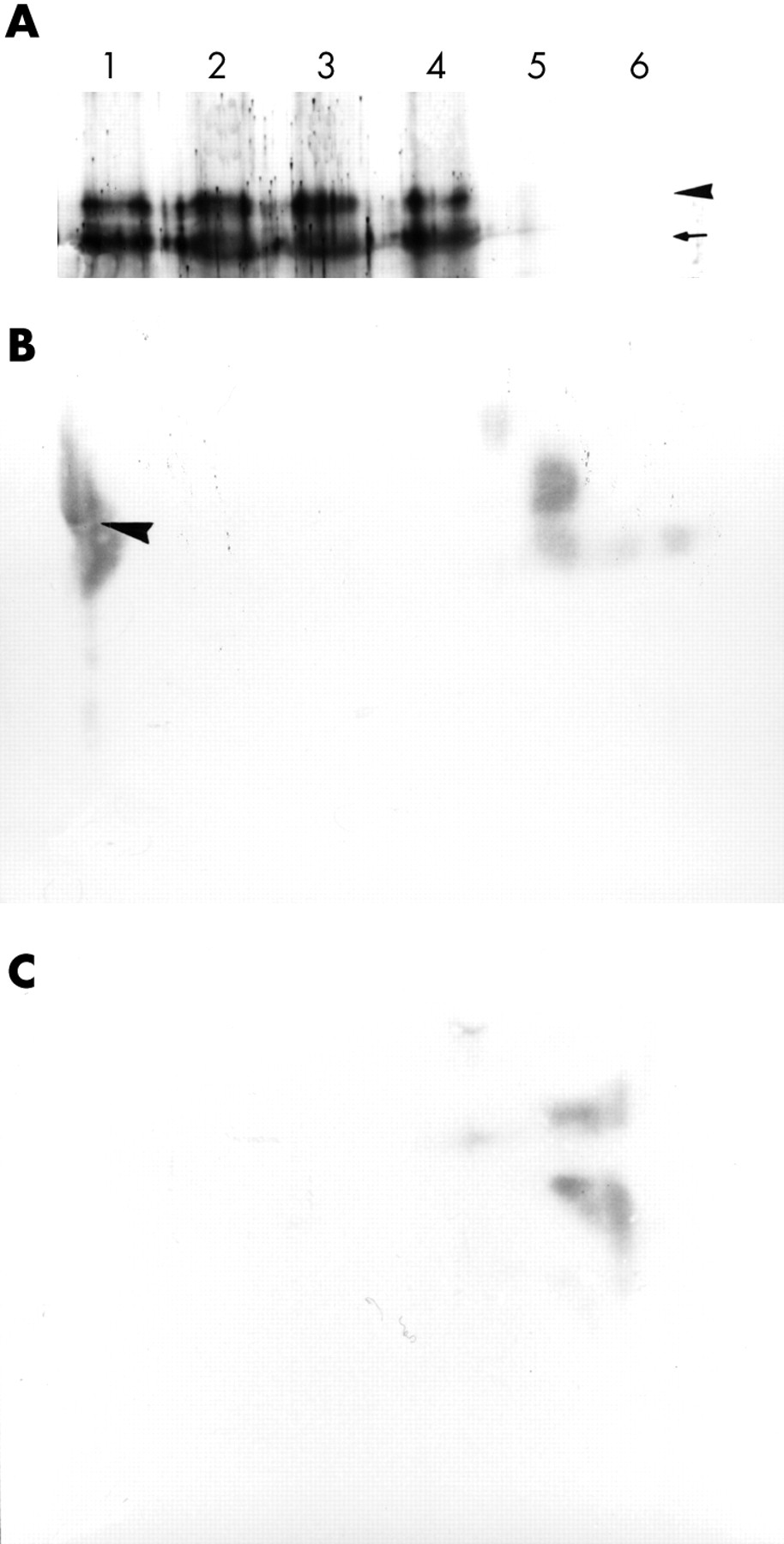
The bands of PCR products. Lane a, 735 bp band (arrow) of human papillomavirus type 16 (HPV-16) in reconstructed plasmid pBR322 using E4a/E5b primer set (positive control). Lane b, 695 bp band (arrowhead ) of HPV-16 transcript using E6a/E7b primer set (16Da cells). Lane c, 735 bp band (arrow) of HPV-16 transcript using E4a/E5b primer set (16Da cells). Lane d, distilled water (negative control). Lane e, 695 bp band (arrowhead) of HPV-16 transcript using E6a/E7b primer set (positive control CaSki cells).
The expression of CDK and cyclin mRNA in HPV transfected cells was similar to that seen in non-HPV transfected cells.
Western blot analysis of involucrin and keratin
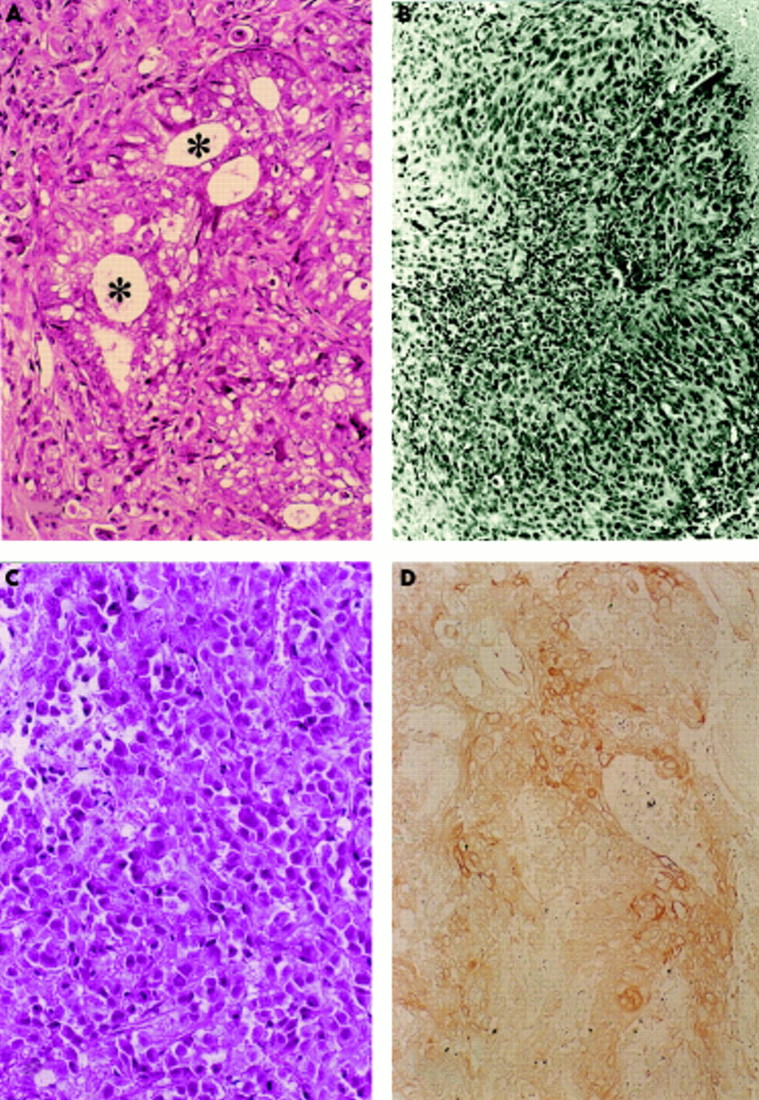
After acrylamide electrophoresis, the purified involucrin was transferred to nitrocellulose membranes, and the bands were visualised using anti-involucrin antibody. Involucrin molecules of 52 kDa and 70 kDa were detected (fig 4A) from all of the HPV-16 transfected cells and control CaSki cell samples.

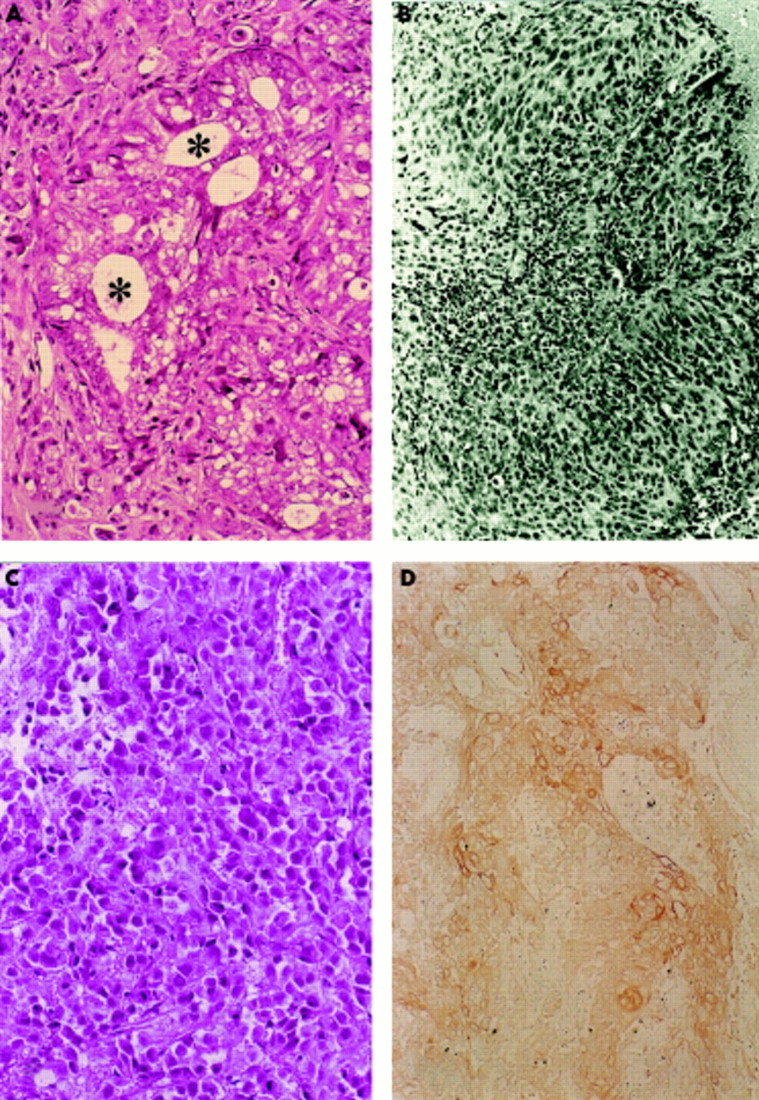
(A) Western blot analysis of 52 kDa (arrow) and 70 kDa (arrowhead) involucrin molecules extracted from 16Da cells. Lane 1, squamous epithelium from the skin of the chest obtained at necropsy; lane 2, squamous epithelium of the oesophagus obtained at necropsy; lane 3, 16Da cells; lane 4, CaSki cells (positive control); lane 5, non-human papillomavirus (HPV) transfected DLD-1 cells; lane 6, distilled water (negative control). (B) Moll’s number 1 keratin (arrowhead) was demonstrated using two dimensional electrophoresis (16Da cells). (C) Control non-HPV transfected DLD-1 cells. Moll’s number 1 keratin was not demonstrated.
Keratin molecules were analysed using two dimensional acrylamide gel electrophoresis. Moll’s number 1 keratin was detected (fig 4B,C) in all of the HPV-16 transfected cells and control CaSki cell extracts. In the 16Da cell (HPV transfected DLD-1 cell), which expressed E2 and E6–E7 fusion gene mRNAs, the expression of involucrin and Moll’s number 1 keratin was higher than that seen in the other HPV transfected clones. Involucrin and Moll’s number 1 keratin were not detected in the non-viral transfected control DLD-1 and PC-14 cells.
Morphological examination
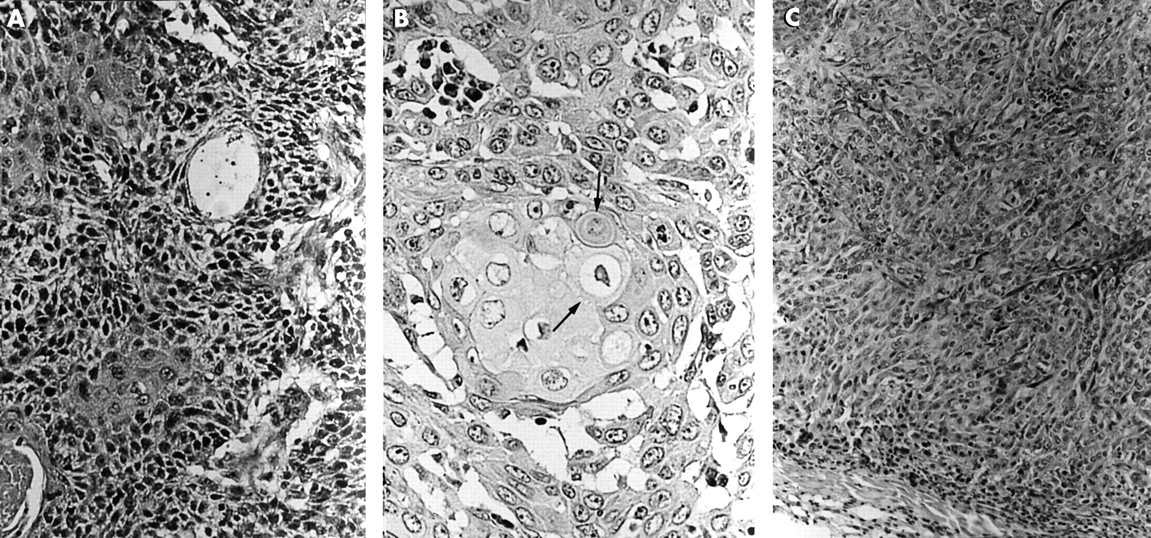
After transfection with HPV, both the DLD-1 and PC-14 cultured cells became larger, with abundant cytoplasm, and were tightly adherent to the dishes. Three to four weeks after injection of these HPV transfected cells into the back of SCID mice tumours formed. The average size of the tumours was about 1.0 cm3. Histopathologically, the tumours were moderately differentiated squamous cell carcinoma, showing abundant cytoplasm with intercellular bridges (figs 5, 6A–C). Immunohistochemical staining for HMK and involucrin in these tumour cells was strong (fig 6D), but mucin was not demonstrated by either alcian blue or PAS staining. Without exception, all of the HPV transfected cells showed squamous metaplasia. The squamous metaplasia was most clearly demonstrated in 16Da cells, which expressed E2 and E6–E7 fusion gene mRNA. The copy number of the transfected virus was not strictly related to the squamous metaplasia. The cells expressing only one of the E2, E6, or E7 genes showed slightly lower HMK and involucrin expression. The L1 and L2 regions were not transcribed in the virus transfected cells, although in the case of CaSki cells L1 region mRNA was detected by RT-PCR. The HPV transfected PC-14 cells also showed squamous metaplasia, with basal cell differentiation, and the number of prickle cells was not so pronounced. In the HPV transfected PC-14 cell clones, no E6–E7 fusion gene mRNA expressing clone was obtained.
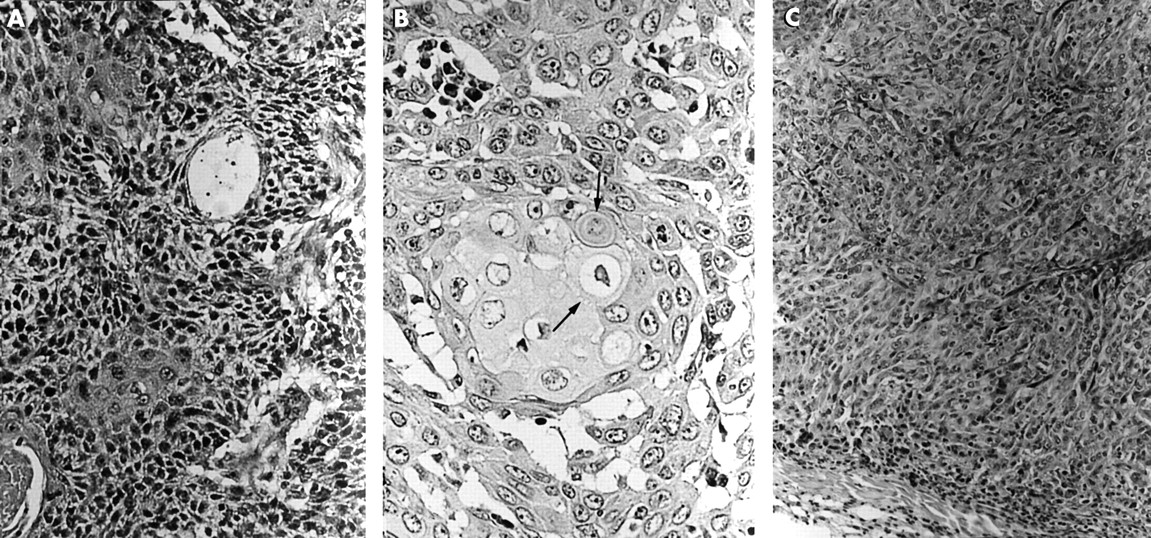
Histological observation of human papillomavirus (HPV) transfected cells. (A) HPV-16 transfected DLD-1 cell (16Da cells). Squamous metaplasia was clearly demonstrated. Haematoxylin and eosin staining; original magnification, ×200. (B) Intercellular bridges (arrows) in the HPV-16 transfected DLD-1 cell (16Da cells); original magnification, ×400. (C) HPV-16 transfected DLD-1 cells (16Db cells) expressing only the E4 and E6 mRNAs. Squamous metaplasia was observed, but prickle cell differentiation was not clear; original magnification, ×200.
(A) Control DLD-1 cells without human papillomavirus (HPV) transfection. The asterisks indicate the tubular structure; original magnification, ×200. (B) HPV-16 transfected PC-14 cells (16Pa cells). Squamous metaplasia was demonstrated, but basal cell-like cells were observed; original magnification, ×200. (C) Control PC-14 cells without HPV transfection. Irregular tubular or trabecular patterns were observed; original magnification, ×200. (D) Demonstration of high molecular weight keratin (HMK). HMK was strongly demonstrated in the HPV-16 transfected DLD-1 cells (16Da cells) with squamous metaplasia; original magnification, ×200.
The tumour cells that expressed the E2 and E6–E7 fusion gene mRNAs expressed HMK and involucrin more strongly than those expressing only one of E2, E4, E5, E6, or E7, although no keratin pearl formation was seen in the HPV transfected cells. Many desmosomes were clearly demonstrated by means of electron microscopy (fig 7). Some cells displayed morphological characteristics of apoptosis, including extensive and pronounced chromatin condensation.

Electron microscopic demonstration of a desmosome. The arrow indicates the desmosome (16Da cells). Original magnification, ×11 000.
Immunohistochemically, the centre of the HPV transfected cell tumour nests (16Da) did not stain for Ki-67 or PCNA, although these molecules were demonstrated in the marginal zone of the nests (data not shown). Using the TUNEL method, large numbers of apoptotic cells were demonstrated in the virus transfected cells (fig 8A,B). Apoptotic cells were frequently seen among cells expressing E2, E6, and E7 mRNAs. However, immunohistochemical staining for the p53 protein was weak in HPV transfected DLD-1 cells, similar to that seen in non-HPV transfected cells.

(A) Apoptotic cells (arrowheads) were demonstrated in 16Da cells using the terminal deoxynucleotidyl transferase mediated dUTP biotin nick end labelling method. Original magnification, ×200. (B) Control DLD-1 cells; apoptotic cells were not demonstrated.
Analysis of the physical state of HPV DNA in virus transfected cells40
In all HPV transformed cells, integrated HPV was demonstrated by in situ hybridisation using Cooper’s criteria20 (fig 9A,B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Non-isotopic in situ hybridisation of human papillomavirus type 16 (HPV-16). Integrated HPV signals (arrowheads) according to Cooper’s criteria were demonstrated in the nuclei of the 16Da cells. All the nuclei of the 16Da cells showed positive signals. Original magnification, ×200. (B) An HPV signal was not demonstrated in the control non-HPV transfected DLD-1 cells. Original magnification, ×200.
The hybridisation pattern of undigested HPV-16 DNA was compared with DNA that has been digested with a “no cut” restriction endonuclease (Hind III or Bgl II), with a “single cut” enzyme (Bam HI), and with a multicut enzyme (Pst I or Eco RI). On Southern analysis the episomal HPV band (7.9 kilobase band after Bam HI digestion) was not demonstrated (data not shown). The HPV transfected cells showed the presence of integrated forms of the virus.
DISCUSSION
HPV transfected cells exhibited low growth in culture, with a high G0–G1 phase cell population, and showed clear “squamous metaplasia” morphologically when they were injected into SCID mice. The induction of HMK (Moll’s10 number 1 keratin), involucrin, and transglutaminase I, which are markers of well differentiated keratinocytes,4,11,12,43–47 was demonstrated in the HPV transfected tumour cells. The tumour cells had abundant cytoplasm and numerous intercellular bridges, and resembled prickle cells or basal cells. It has been reported previously2 that squamous metaplasia is more clearly demonstrated in HPV-16 transfected DLD-1 cells (16Da) than other HPV type transfected cells. However, in the case of PC-14 cells, “basal cell differentiation” was frequently observed, and prickle cells were less frequently found. It is thought that the poorly differentiated characteristics of the PC-14 cells, which were established from poorly differentiated adenocarcinoma of the lung, and the expression pattern of HPV mRNA might influence the differentiation of the metaplastic cells. In our present study, no PC-14 cell clone was obtained that expressed HPV E2 and E6–E7 fusion gene mRNA simultaneously.
“The combined expression of the E2 and E6–E7 fusion gene products might be a factor for squamous metaplasia”
Among the HPV transfected cells, the HPV-16 transfected DLD-1 cell (16Da), which expressed E2 and E6–E7 fusion gene mRNA, showed squamous metaplasia particularly clearly. However, keratin pearl formation was not demonstrated in our present experiments and it is possible that keratinising lesions may be formed by further differentiation factors,4,43,48,49 including calcium ions or a growth factor.9,20,40 Choo et al reported that calcium increased the differentiation of HPV-16 positive CaSki cells.43 The influence of different culture conditions needs to be examined further.
It has been reported that HPV genomes are transcribed into multiple and differentially spliced mRNAs.6,40,41,45,50–52 Transcription is subject to complex control by viral and cell functions. In the present HPV transfected cells, clear squamous metaplasia was seen in the clones that expressed the E2 and E6–E7 fusion genes simultaneously. In these cases, in addition to fusion transcripts of E6–E7, small amounts of E2–E6 and E4–E5 were also detected, but L1 and L2 mRNA was not. The expression of E4 and E5 was variable among the cell lines and was not thought to correlate with squamous metaplasia. In our present cell lines, only cells expressing single E6 or E7 transcripts did not produce pronounced squamous metaplasia. Therefore, the combined expression of the E2 and E6–E7 fusion gene products might be a factor for squamous metaplasia. Products from the E2 gene region have been implicated previously in the regulation of HPV gene transcription.41,50,53 Integration of the viral genome generally results in the disruption of the E2 open reading frame, which encodes a transactivator of viral transcription.42 However, DLD-1 clone 16Da, which expressed E2 mRNA without disruption, showed clear squamous metaplasia. The transactivator E2 protein and E6–E7 fusion protein might be important for HPV induced squamous metaplasia, although the amount of E6–E7 and E2 transcripts did not influence the development of squamous metaplasia. L1 and L2 were not necessary for the development of squamous metaplasia. Furthermore, a pronounced degree of apoptosis was demonstrated in the 16Da cell clone. E6 and E7 were considered necessary for apoptosis, as reported elsewhere.45,54,55 In 16Da cells, DNase γ and AIF were strongly expressed, although caspase 3, caspase 9, and DNase I mRNA values were similar among all HPV transfected and non-transfected control cells. The Fas ligand was not expressed, but a small amount of Fas mRNA was detected in a few clones of HPV transfected DLD-1 and PC-14 cells. The bcl-2 and bax mRNA values were unchanged among all cells. It was thus considered that the strong expression of DNase γ in 16Da cells might depend on a unique pathway, different to the caspase 3 and caspase 9 associated route. AIF was strongly expressed, but caspase 9 expression was lower than that of caspase 8. Furthermore, in one of the HPV-16 transfected cells DNase γ was expressed, but caspase 8 was not detected. The expression of caspase 8 was variable among the cell clones, and small amounts of it were also detected in the non-virus transfected control DLD-1 and PC-14 cells.
It is well known that the cervical epithelium infected with HPV develops intraepithelial neoplasia. Most of the intraepithelial neoplasia of the cervix occurs in the transitional zone, which is thought to be composed of squamous metaplastic cells. Sun et al reported that cultured normal endocervical epithelial cells infected with HPV-16 formed an epithelium resembling squamous metaplasia.47 However, neither detailed examinations of HPV transcripts nor molecular biology were performed. Merrick and colleagues56 also reported the altered expression of differentiation markers in the squamous epithelial cells immortalised by HPV-16 and HPV-18. In our report, we used HPV-16 infected CaSki cells as a control. HeLa cells infected with HPV-18 should also be analysed for comparison. Furthermore, in the case of uterine endometrioid carcinoma, it is well known that squamous cell metaplasia or “squamous differentiation” is frequent.13 Therefore HPV infection in endometrioid carcinoma should also be examined, and indeed cultured endometrioid carcinoma cells are another appropriate candidate for HPV induced experimental squamous metaplasia. But squamous metaplasia is a change in the normal differentiation process of the progenitor cell through a different pathway of differentiation. In our study, cultured cancer cells were used, but normal tissue—for example, bronchial epithelium—should be used for an in vivo experiment. HPV infection was demonstrated with adenocarcinoma cells as reported previously,3 but HPV infected adenocarcinoma cells showed an increased area of cytoplasm and developed into adenosquamous carcinoma. The HPV integration site in chromosomes varies from case to case, although Cannizzaro and colleagues57,58 have reported integration at the chromosome 3p fragile site. It is thought that HPV infects epithelial cells and that the viral gene, especially the E6, E7 and E2 products,59 induces the abrogation of the cell cycle,46,60 and that in this process squamous metaplasia is induced, and dysplasia and neoplasia develop through further gene alterations. Recently Graham and colleagues49 have indeed reported that additional genetic alterations in HPV infected cells are thought to be required for the development of a carcinogenic genotype.
Take home messages
-
Human papillomavirus (HP) transfection into adenocarcinoma cells induced clear squamous metaplasia
-
Squamous metaplasia was particularly clear in one of the HPV transfected cell clones, which expressed E2 and E6–E7 fusion gene mRNA, and apoptosis was also demonstrated
-
Further studies are needed to elucidate which sequences in the viral genome—E2, E6, or E7—are the most important factors for squamous metaplasia
“A high degree of apoptosis was demonstrated in those transfected cell clones that simultaneously expressed both E2 and E6–E7 fusion gene mRNA, but not L region mRNA”
Furthermore, it is well known that bovine papillomavirus (BPV) E2 proteins can repress E6 and E7 expression.61,62 Although overexpression of the BPV E2 protein decreases the transcription of HPV-16 from the P97 promoter, under identical conditions, the HPV-16 E2 protein has also been shown to activate the transcription of HPV from the same promoter.63 Furthermore, Sanchez-Perez and colleagues55 and Bouvard and colleagues63 have reported that the HPV-16 E2 protein increases E6 and E7 mRNA values, and that an increase in the E7 protein concentration induces apoptotic cell death.45,55 Reintroduction of HPV-16 E2 protein into an HPV-16 transformed cervical carcinoma cell line (SiHa cell), which contains the integrated HPV 16 DNA with a disrupted E2 gene, results in a decrease in growth rate.55 Furthermore, the E6 and E7 genes are reported to induce changes in the expression of cell growth proteins, and also in involucrin expression.38 In our present studies, apoptotic cells were demonstrated by HPV genome transfection. A high degree of apoptosis was demonstrated in those transfected cell clones that simultaneously expressed both E2 and E6–E7 fusion gene mRNA, but not L region mRNA.
Further studies are needed to elucidate which sequences in the viral genome—E2, E6, or E7—are the most important factors for squamous metaplasia.