Article Text

Abstract
Background Chronic myeloproliferative disorders (MPDs) are clonal haematopoietic stem cell malignancies characterised by an accumulation of mature myeloid cells in bone marrow and peripheral blood. Deregulation of the apoptotic machinery may be associated with MPD physiopathology.
Aims To evaluate expression of death receptors' family members, mononuclear cell apoptosis resistance, and JAK2 allele burden.
Subjects and Methods Bone marrow haematopoietic progenitor CD34 cells were separated using the Ficoll-hypaque protocol followed by the Miltenyi CD34 isolation kit, and peripheral blood leukocytes were separated by the Haes-Steril method. Total RNA was extracted by the Trizol method, the High Capacity Kit was used to synthesise cDNA, and real-time PCR was performed using SybrGreen in ABIPrism 7500 equipment. The results of gene expression quantification are given as 2−ΔΔCt. The JAK2 V617F mutation was detected by real-time allelic discrimination PCR assay. Peripheral blood mononuclear cells (PBMCs) were isolated by the Ficoll-hypaque protocol and cultured in the presence of apoptosis inducers.
Results In CD34 cells, there was mRNA overexpression for fas, faim and c-flip in polycythaemia vera (PV), essential thrombocythaemia (ET) and primary myelofibrosis (PMF), as well as fasl in PMF, and dr4 levels were increased in ET. In leukocytes, fas, c-flip and trail levels were increased in PV, and dr5 expression was decreased in ET. There was an association between dr5 and fasl expression and JAK2V617F mutation. PBMCs from patients with PV, ET or PMF showed resistance to apoptosis inducers.
Conclusions The results indicate deregulation of apoptosis gene expression, which may be associated with MPD pathogenesis leading to accumulation of myeloid cells in MPDs.
- Apoptosis
- myelofibrosis
- myeloproliferative disease
- polycythaemia
- thrombocythaemia
Statistics from Altmetric.com
Introduction
Myeloproliferative disorders (MPDs) are clonal haematopoietic stem cell malignancies characterised primarily by hypercellular bone marrow, proliferation of cells from one or several myeloid lineages, and haematopoietic progenitor independence of, or hypersensitivity to, cytokines.1–3
According to the World Health Organization (WHO), myeloproliferative neoplasms can be divided into two groups: classic MPDs, comprising chronic myelogenous leukaemia (CML), polycythaemia vera (PV), essential thrombocythaemia (ET) and primary myelofibrosis (PMF); and less common MPDs, such as chronic neutrophilic leukaemia.4
MPDs are heterogeneous in their molecular aspects and natural course, but their physiopathology often involves deregulation of tyrosine kinases, such as that of Bcr-Abl in the pathogenesis of CML.1 5
In 2005, several independent groups reported in patients with MPD a somatic mutation in the Janus kinase JAK2, which has a key role in the signal transduction pathway of various haematopoietic growth factor receptors.2 6–9 The point mutation that led to valine-to-phenylalanine substitution at polypeptide position 617 is designated JAK2V617F and is responsible for constitutive kinase activity.10–14
JAK2V617F mutation has been reported in approximately half of all patients with PMF and ET and in nearly all of those with PV.2 6–8 15–17 Increased JAK2V617F allele burden has been associated with upregulated expression of downstream JAK2 target genes, such as polycythaemia rubra vera 1 (PRV-1).6 16 17 There are also indications that patient mutation status correlates with prognosis and treatment response.18–21
Regardless of the recent advances in MPD diagnosis and description of molecular markers, clearer reports of the mechanisms involved in the physiopathology of MPDs are needed to identify new markers of diagnostic and prognostic molecular targets for therapy. In this context, we can hypothesise that deregulation of apoptosis contributes to the myeloaccumulation seen in these disorders, since there is a constant need to control the numbers of haematopoietic cells because of the great potential for intrinsic proliferation and cell turnover.22–24 The literature data reinforce our hypothesis, since deregulation of the apoptosis process has been reported in various myeloid haematological neoplasias such as chronic neutrophilic leukaemia,25 myelodysplastic syndromes,26 CML27 and in MPD chromosome Ph negative.28 29 In myelodysplastic syndromes, it has been described as an altered expression of genes related to the intrinsic apoptosis pathway.26 In CML, the cells are resistant to apoptosis due to the kinase Bcr-Abl, and overexpression of anti-apoptotic genes has been described.27 In MPD chromosome Ph negative, little is known about apoptosis-related gene expression. Zeuner et al28 in PV found c-flip overexpression, and Ciurea et al29 described a different development of megakaryocyte apoptosis and overexpression of anti-apoptotic protein Bcl-xL in PMF. Laubach and collaborators30 reported that PV-deregulated erythropoiesis is linked to erythropoietin hypersensitivity and apoptosis resistance of erythroid precursor cells.
Taken together, all this information from the literature indicates that, in most cases, apoptosis deregulation is associated with haematological neoplasia pathogenesis.25–31
Two major apoptotic pathways have been well characterised: one is activated by changes in mitochondrial permeability (intrinsic pathway); the other is triggered by cell surface death receptors engaging their specific ligands (extrinsic pathway).32 Activation of the death receptor pathway, which primarily includes CD95 (APO-1/Fas) or TRAIL receptors (DR4 and DR5) and their ligands (FasL and TRAIL, respectively), likewise the anti-apoptotic molecules FAIM and C-FLIP, results in recruitment of adaptor molecules and caspase-8 to form the death-inducing signalling complex, thereby inducing caspase-3 activation and apoptosis.24 33
The aim of this study was to quantify expression of the genes fas, fasl, faim, c-flip, dr4, dr5 and trail, all of which participate in mechanisms that regulate the extrinsic pathway, in CD34 cells and leukocytes from patients with MPDs. We also attempted to detect mononuclear cell susceptibility to death stimulated by apoptosis inducers and to correlate the results with JAK2V617F allele burden.
Subjects and methods
Patients and controls
In this study, bone marrow and peripheral blood samples from 26 patients with ET, 12 with PV, and 11 with PMF were evaluated. There were 17 men and 32 women, mean age was 61.1 years (range, 35–83), and 43 patients were white and six were black (online supplementary table 1). All of the patients selected had been diagnosed according to WHO criteria.4 The controls comprised bone marrow samples collected from 23 healthy subjects (12 male, 11 female; mean age 35.9 years (range 14–54)) and peripheral blood samples from 30 subjects (11 male, 19 female; mean age 54.5 years (range 31–80)). Patients were selected from those treated at the Brigadeiro Hospital as well as from the Hospital das Clínicas of the University of São Paulo at Ribeirão Preto School of Medicine, state of São Paulo, Brazil. Healthy subjects were recruited from the University of São Paulo at Ribeirão Preto School of Pharmaceutical Sciences, whereas those who provided bone marrow samples were recruited from among marrow donors at the Bone Marrow Transplantation Unit of the Hospital das Clínicas of the University of São Paulo. There is a significant age difference between bone marrow controls and the patient population because the local ethics committees would only permit bone marrow samples to be obtained from bone marrow donors during the bone marrow transplantation cell collection procedure.
Isolation of peripheral blood mononuclear cells (PBMCs) and leukocytes
PBMCs were isolated using the Ficoll-hypaque density gradient protocol (Histopaque; Sigma-Aldrich, St Louis, Missouri, USA) according to the manufacturer's instructions. The PBMCs isolated were used to assess lymphocyte apoptosis assays.
Leukocytes were obtained from the peripheral blood of patients and controls using Haes-Steril reagent (Voluven; Frasenius Kabi, Campinas, Brazil).
CD34 haematopoietic stem cell separation
Haematopoietic progenitor CD34 cells collected from patients and controls were separated from bone marrow mononuclear cells using the MidiMacs CD34+ Isolation Kit (MACS; Miltenyi Biotec, Bergisch Gladbach, Germany) according to the manufacturer's protocol. CD34 purity was determined by flow cytometry (FACSort; Becton Dickinson, San Jose, California, USA). CD34 cells with ≥80% purity were used in the detection of gene expression by quantitative real-time PCR.
Total RNA extraction, cDNA synthesis and real-time PCR
Total RNA was extracted from leukocytes and CD34 cells using Trizol methods (Invitrogen Life Technologies, Carlsbad, California, USA) as described by the manufacturer. RNA concentration and purity was determined spectrophotometrically by measuring fluorescence at 260 nm and 280 nm. RNA (1 μg) was reverse-transcribed to cDNA using the High Capacity cDNA Archive Kit (Applied Biosystems, Foster City, California, USA), also according to the manufacturer's instructions. cDNA (1 μl of cDNA diluted 1:3) was applied in the real-time PCR assay to determine the expression of c-flip, dr4, dr5, fas, fasl, faim and trail. Gene expression was quantified using the SYBR Green PCR Master Mix Kit (Applied Biosystems) and specific primers (Invitrogen Life Technologies) (online supplementary table 2) on the 7500 Real Time PCR System (Applied Biosystems). Real-time PCR conditions were: 50°C for 2 min; 95°C for 10 min; and 50 cycles at 95°C for 15 s; 54 at 60°C (specific for target gene) for 25 s; and 72°C for 34 s. The annealing temperatures of each set of primers are also shown in supplementary table 1. β-actin and GAPDH genes were used as endogenous controls (housekeeping genes). The amount of mRNA for each sample was normalised using the geometric average of the housekeeping genes. All reactions were duplicated, and the relative expression levels were calculated using the 2−ΔΔCt method34 with the mean of the control group used as calibrator.
Detection of JAK2V617F mutation and determination of JAK2V617 allele burden
The JAK2V617F mutation was detected in a reference laboratory (Fleury Medicine and Health, São Paulo, SP, Brazil) by real-time allelic discrimination PCR assay. The assay used two fluorogenic, minor groove binding probes to detect the G→T mutation in exon 14 of JAK2 gene. Sequences of primers (forward primer, 5′-GCAGCAAGTATGATGAGCAAGCT-3′; reverse primer, 5′-GGCATTAGAAAGCCTGTAGTTTTACTTAC-3′) and minor groove binding probes were designed by using Primer Express software, version 1.5 (Applied Biosystems). The hybridisation probes were designed with different dyes to allow single-tube allelic discrimination. One probe with the reporter FAM (6-carboxyfluorescein) was targeted to the wild-type allele (5′-6-FAM-TGGAGTATGTGTCTGTGGA-3′) and the other probe with the reporter VIC (6-carboxyrhodamine 6G) was targeted to the mutant allele (5′-VIC-TGGAGTATGTTTCTGTGGAG-3′). Primers were synthesised by IDT (Integrated DNA Technologies, Coralville, Iowa, USA) and probes were synthesised by Applied Biosystems. Amplification by real-time PCR was performed using TaqMan Universal Master Mix (Applied Biosystems). Thermal cycling conditions consisted of 50°C for 2 min, 45 cycles of 95°C for 15 s and 60°C for 1 min on the Rotor-Gene 3000 (Corbett Research, Sydney, Australia). Each PCR assay included a control without added DNA (no-template control).
The allele burden was estimated based on the normalised fluorescent signal of the mutant allele and wild-type allele of real-time PCR and reported as a percentage of JAK2V617F mutation or negative.
Assessment of mononuclear cell apoptosis
Mononuclear cells were cultured in complete RPMI 1640 medium (Sigma) supplemented with 10 mM Hepes, 200 mM l-glutamine, 100 units/ml penicillin, 100 μg/ml streptomycin and 10% fetal bovine serum at 37°C in an atmosphere of 5% CO2 for 12 h in the presence of apoptosis inducers: actinomycin (ACT-D, 1 μM; Sigma); etoposide (VP16, 25 μM; Bristol-Meyers-Squibb, Mayaguez, Puerto Rico); teniposide (VM26, 25 μM; Bristol-Meyers-Squibb); cytarabine (ARA-C, 25 μM; Pfizer, Milano, Italy); staurosporine (STS, 0.5 μM; Sigma-Aldrich).
Cell apoptosis was quantified by flow cytometry, using the annexin-V/fluorescein isothiocyanate (FITC) technique. Mononuclear cells were recovered from culture medium by centrifugation at 240 x g for 10 min at 4°C, washed once with annexin buffer (10 mM Hepes, pH 7.4; 150 mM NaCl; 5 mM KCl; 1 mM MgCl2; 1.8 mM CaCl2) and incubated for 20 min in the dark with 100 μl annexin-V/FITC. Subsequently, 40 μl propidium iodide (PI) solution (50 μg/ml) was added to each tube, and the contents were analysed by flow cytometry. Five thousand mononuclear cells were acquired using a FACS Canto Flow Cytometer (Becton-Dickinson) and analysed by dot-plot using Diva 6.0 Software (Becton-Dickinson). The results are given as a percentage of the apoptotic cells (mononuclear cells positive for annexin-V and PI negative staining). A representative FACS plot of annexin/PI staining showing the full gating strategy on mononuclear cells (forward scatter and side scatter gate) and the dot-plot analysis of negative control and mononuclear cells stimulated by teniposide (VM26) are shown in figure 1A–C.

(A) Dot-plot diagram of gate in control mononuclear cells; (B) dot-plot diagram of fluorescein isothiocyanate (FITC)/annexin-V/propidium iodide (PI) flow cytometry of control mononuclear cells without treatment; (C) dot-plot diagram of FITC/annexin-V/PI flow cytometry of control mononuclear cells treated with 25 μM teniposide. In (B) and (C), the lower right quadrants show the viable cells, which exclude PI and are negative for FITC/annexin-V binding. The upper right quadrants contain the non-viable cells, necrotic cells, positive for FITC/annexin binding and for PI uptake. The lower right quadrants represent apoptotic cells, FITC/annexin-V positive and PI negative. FSC: Forward scatter; SSC: side scatter; FL1: fluorescence channel 1; FL3: fluorescence channel 3.
Although it is controversial, many authors35–38 have reported lymphoid lineage involvement in PV, ET and PMF. Thus, we chose mononuclear cells to perform the assays of apoptosis measurement because the mononuclear cell isolation protocol produces a cell population with high purity, and FACS analysis well discriminates between lymphocytes and monocytes stimulated by apoptotic inducers.
Statistical analysis
Data related to gene expression and percentages of apoptotic cells from patients and controls were compared by the non-parametric Mann–Whitney test. To draw correlations between JAK2V617F allele burden and gene expression, we used the non-parametric Spearman test. Statistical analyses were carried out using Prism Software V5.0. p≤0.05 was considered significant.
Results
fas, fasl, dr4, faim and c-flip expression is elevated in CD34 cells from patients with MPDs
As can be seen in figure 2A, fas expression was significantly higher in the PV, ET and PMF groups than in the control group (p<0.0001, p=0.0013 and p=0.0004, respectively), and there was significant difference between the control group and PMF group in terms of CD34 fasl mRNA levels (p=0.0002, figure 2B). The levels of trail and dr5 in CD34 cells were comparable between patients and controls (p >0.05) (online supplementary figure 1A,B). In contrast, dr4 expression was increased in the ET group (p=0.0079; figure 2C).

Gene expression in bone marrow CD34 cells of control, polycythaemia vera (PV), essential thrombocythaemia (ET) and primary myelofibrosis (PMF) groups: (A) fas (median: control, 1.80; PV, 13.16; ET, 5.52; PMF, 7.21); (B) fasl (median: control, 1.13; PV, 11.16; ET, 4.46; PMF, 213.1); (C) dr4 (median: control, 1.54; PV, 2.49; ET, 5.20; PMF, 8.43); (D) faim (median: control, 0.82; PV, 12.95; ET, 26.03; PMF, 23.56); (E) c-flip (median: control, 0.78; PV, 7.22; ET, 1.93; PMF, 8.37). All differences are p>0.05 unless otherwise noted.
Expression of the anti-apoptotic gene faim was also elevated in the three groups in comparison with the control group (p=0.0048, p=0.0002, p=0.0076), as shown in figure 2D, and c-flip expression was also elevated in the PV, ET and PMF groups in comparison with the control group (p=0.0228, p=0.0121 and p=0.0026, respectively; figure 2E).
Analysing gene expression among MPD groups, we found a significant difference between the ET and PMF groups in terms of fasl expression (p=0.0012, figure 2B).
It is worth emphasising here that there is no difference in apoptosis-related gene expression (p>0.05) with age (range 14–54 years, median 37.0) within the normal population (bone marrow donors) (data not shown).
Expression of fas, trail, dr5, c-flip and faim is deregulated in leukocytes of patients with MPDs
Expression of fas was higher in the PV group than in the control group (p=0.0013; figure 3A), and fasl expression was not significantly different between MPD groups and the control group (p>0.05) (online supplementary figure 1C). In addition, we found that trail mRNA concentration was higher in PV leukocytes than control group leukocytes (p=0.0321, figure 3B), and dr5 expression was lower in ET leukocytes (p=0.0387, figure 3C). dr4 expression was not significantly different between MPD groups and the control group (p>0.05) (online supplementary figure 1D).

Gene expression in peripheral leukocytes of control, polycythaemia vera (PV), essential thrombocythaemia (ET) and primary myelofibrosis (PMF) groups: (A) fas (median: control, 0.60; PV, 4.38; ET, 0.51; PMF, 1.11); (B) trail (median: control, 1.024; PV, 2.25; ET, 0.67; PMF, 0.89); (C) dr5 (median: control, 0.92; PV, 1.40; ET, 0.52; PMF, 0.77); (D) faim (median: control, 1.18; PV, 1.49; ET, 1.37; PMF, 2.05); (E) c-flip (median: control, 0.81; PV, 9.71; ET, 1.32; PMF, 3.97). All differences are p>0.05 unless otherwise noted.
Expression of the anti-apoptotic genes faim and c-flip was increased in the PMF (p=0.0040) and PV (p=0.0043) group, respectively (figure 3D,E).
Analysing MPD leucocyte gene expression, we found a significant difference between PV and ET groups in terms of fas expression (p<0.0001, figure 3A) and trail (p=0.0033, figure 3B) and between PV and PMF groups in terms of fas and trail expression (p=0.0053 and p=0.0123, respectively, figure 3A,B). We also detected that fas expression was elevated in PMF patients in comparison to ET patients (p=0.0326, figure 3A).
Correlation between gene expression and JAK2V617F mutation
There was a correlation between the JAK2V617F allele burden and fasl expression (r=−0.5053; p=0.0469; 95% CI=−0.8423 to 0.1159) in CD34 and between JAK2V617F allele burden and dr5 expression (r=0.6246; p=0.0150; 95% CI=0.05985 to 0.8865) in leukocytes from PV patients (figure 4A,B, respectively).

Correlation of (A) fasl and (B) dr5 expression in polycythaemia vera (PV) patients with the JAK2V617F allele burden.
There was no association between apoptosis-related gene expression and allele burden in ET and PMF patients. No differences were observed in level of apoptosis-related gene expression between JAK2V617F positive and negative ET and PMF patients.
Mononuclear cells of MPD patients are resistant to apoptosis inducers
Mononuclear cell resistance to 1 μM ACT-D and 25 μM VP16 was higher in the PV (p=0.03, p=0.03, respectively), ET (p<0.0001, zp=0.01, respectively) and PMF (p<0.0001, p<0.0001, respectively) groups than in the control group (figure 5A,B). Cells obtained from ET and PMF patients were more resistant to apoptosis induced by 25 μM VM26 than were those obtained from controls (p=0.005 and p=0.001, respectively; figure 5C). In comparison with the values obtained for controls, the resistance of apoptotic cells to 25 μM ARA-C was higher in PV, ET and PMF group cells (p=0.031, p=0.033 and p=0.013, respectively; figure 5D), as was that induced by 5 μM STS in ET and PMF group cells (p=0.0002 and p<0.0001, respectively, figure 5E). Among MPD groups, we detected a difference in mononuclear cell susceptibility to: 1 μM ACT comparing PV and ET groups (p=0.004), PV and PMF groups (p=0.002) and ET and PMF groups (p=0.05) (figure 5A); 25 μM VP16 between PV and PMF groups (p=0.014) and between ET and PMF groups (p=0.004) (figure 5B); 25 μM VM26 comparing PV and PMF groups (p=0.007) and comparing ET and PMF groups (p=0.047) (figure 5C); and 0.5 μM STS between PV and ET groups (p=0.002), between PV and PMF groups (p=0.0009) and between ET and PMF groups (p=0.022) (figure 5E).

Percentage of mononuclear cell apoptosis induced by drugs and detected by annexin-V/fluorescein isothiocyanate (FITC) in control, polycythaemia vera (PV), essential thrombocythaemia (ET) and primary myelofibrosis (PMF) groups: (A) 1 μM ACT-D (median: control, 35.10; PV, 25.50; ET, 13.00; PMF, 5.25); (B) 25 μM VP16 (median: control, 21.41; PV, 12.25; ET, 15.70; PMF, 4.90); (C) 25 μM VM26 (median: control, 26.80; PV, 21.00; ET, 19.60; PMF, 7.70); (D) 25 μM ARA-C (median: control, 9.10; PV, 5.40; ET, 6.35; PMF, 1.90); (E) 0.5 μM STS (median: control, 36.05; PV, 30.35; ET, 15.80; PMF, 5.50). All differences are p>0.05 unless otherwise noted. The box and whisker graphs show the maximum, median and minimum of the mononuclear cell apoptosis percentage.
Discussion
MPDs are characterised by increased numbers of myeloid cells, a lineage that typically predominates. The mechanisms whereby myeloproliferation, disease progression and development occur in MPDs remain unknown. Abnormal expression of apoptosis regulatory genes has been described in oncological and haematological diseases, and associations between regulatory gene expression and the pathogenesis of such diseases have been demonstrated.26 28 39
Thus, we investigated here the expression of the genes fas, fasl, faim, c-flip, dr4, dr5 and trail in CD34 haematopoietic stem cells and leukocytes from patients with MPDs. We detected a potential correlation between apoptosis-related gene expression with JAK2V617F mutation and mononuclear cell apoptosis.
It is noteworthy that we found alterations in Fas system-related gene expression in CD34 cells and leukocytes from patients with MPDs. It is well known that the Fas/FasL system is critically involved in haematopoietic cell survival and apoptosis.24 33 40 Under physiological conditions, CD34 bone marrow cells express Fas at lower levels; during haematopoietic cell differentiation, the receptor is expressed in proliferating and differentiating progenitors, serving as a negative regulator of distal differentiation in all lineages.41–43 FAIM, the expression of which is also deregulated in MPD patients evaluated here, is a Fas antagonist that has been reported to be upregulated in resistant B cells and functions as an inhibitor of Fas-induced cell death. FAIM is highly conserved in evolution and is broadly expressed in many tissues.44–47
Regarding c-flip expression, our data corroborate data in the literature on PV and ET.28 48 Zeuner et al28 showed an increase in FLIPshort expression in PV erythroid precursor cells. These authors found that JAK2V617F mutation in PV erythroblasts obtained in vitro was associated with cytokine-independent activation of the ERK/MAPK and PI3K pathways and deregulated expression of c-flip. They also showed that one patient in three with wild-type JAK2 was resistant to death receptor activation and had extremely high levels of c-flip, indicating that c-flip overexpression may also contribute to death receptor resistance in PV patients with wild-type JAK2.28
Tenedini et al48 described overexpression of c-flip in a global gene expression study of normal and malignant megakaryocytic cells from ET patients. They concluded that there is an imbalance between proliferation and apoptosis in malignant ET megakaryocytopoiesis.48 We also evaluated the ligand TRAIL, a member of the tumour necrosis factor superfamily of genes, and expression of its two death receptors, DR4 and DR5. The true physiological role of the TRAIL molecule remains unclear. However, various studies have shown that it is a potent inductor of apoptosis in a wide range of cancer cell lines, although not in many normal cells.30 49–51
In the present study, we did not find any alteration in trail expression in CD34 cells, but it was increased in leukocytes from MPD patients. Deregulated expression of death receptors dr4 and dr5 in CD34 cells and leukocytes from ET was also observed. Despite the fact that there are few data on apoptotic gene expression in MPD patients, these findings are in agreement with our hypothesis that apoptosis is deregulated in Bcr-Abl-negative patients with MPD. Besides, these molecules may be used in the future as molecular markers or prognostic factors for these diseases, since dr4 expression has been shown to correlate positively with tumour grade in patients with breast cancer who have invasive ductal carcinoma,52 and it has been shown that there is a significant association between high dr5 expression and reduced overall survival.53 Furthermore, TRAIL therapy has been used in the treatment of neoplasia, and resistance to this treatment due to mechanisms such as changes in the expression of the anti-apoptotic proteins c-flip and mcl-154 has been demonstrated.55 56
It is well known that the clinical course is less favourable among JAK2V617F-positive patients, and a high percentage of JAK2V617F mutations have been found to be associated with proliferation of clonogenic progenitors independently of exogenous cytokine stimulation.57 It has been speculated that constitutive activation of JAK2 correlates with changes in gene expression, since JAK2V617F activates the ERK/MAPK and PI3K pathways, as well as the STAT5 pathway, resulting in sustained phosphorylation of these molecules and cell growth in the absence of the appropriate stimulus.2 This would also protect cells from death receptor-induced apoptosis through multiple mechanisms, including ‘upmodulation’ of c-flip.28 58–60 Our results suggest a relation between this molecular abnormality and fasl and dr5 expression in PV patients. However, as we analysed seven genes and three diseases in this study, and the significance of the correlation of JAK2V617F allele burden and fasl expression was weak, we cannot confirm that the deregulated expression of the apoptosis-related genes analysed in this study is linked to JAK2V617F mutation.
We also assessed mononuclear cell apoptosis in MPD patients by using apoptosis inducers. The results suggest that the mononuclear cells of MPD patients were resistant to most of the drugs used in the assay, whereas those of the control group were not. The apoptosis inducers used in this study had different mechanisms of action for activating the intrinsic and extrinsic apoptosis pathways. For example, ACT-D inhibits DNA synthesis, triggering the intrinsic pathway, and also decreases expression of the anti-apoptotic gene c-flip, as well as amplifying fasl and TRAIL-mediated apoptosis.61 The percentage of apoptosis was lowest in patients with PMF, indicating that the level of resistance was highest in those patients.
c-flip overexpression in MPDs may block the apoptosis process by inhibiting procaspase-8 activation at the death-inducing signalling complexes by blocking its processing. Inhibition of caspase-8 cleavage leads to impairment of the activation of downstream effector caspases, hampering propagation of the apoptotic signal. faim upregulation also contributes to cell resistance to apoptosis in MPDs, since this molecule is a mediator of Fas resistance. FAIM plays a role in modulating Fas-mediated apoptosis and may act by influencing expression of c-FLIP and regulating the physical binding of caspase-8 to Fas.62 Deregulation of fasl, fas and dr4 expression in the MPD patients evaluated may have also interfered with cell sensitivity to apoptosis; however, faim and c-flip negatively regulated activation of apoptosis pathway downstream receptors.
Taken together, our results indicate that MPD results in deregulation of the expression of genes that regulate the extrinsic pathway of the apoptotic mechanism (figure 6). To our knowledge, this is the first study to simultaneously evaluate the expression of these specific anti- and pro-apoptotic death receptor pathway genes in Philadelphia-negative MPD patients, associating the results with apoptosis function and JAK2 allele burden. The data obtained in this investigation support the idea that deregulation of apoptosis may contribute, at least in part, to MPD pathogenesis.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
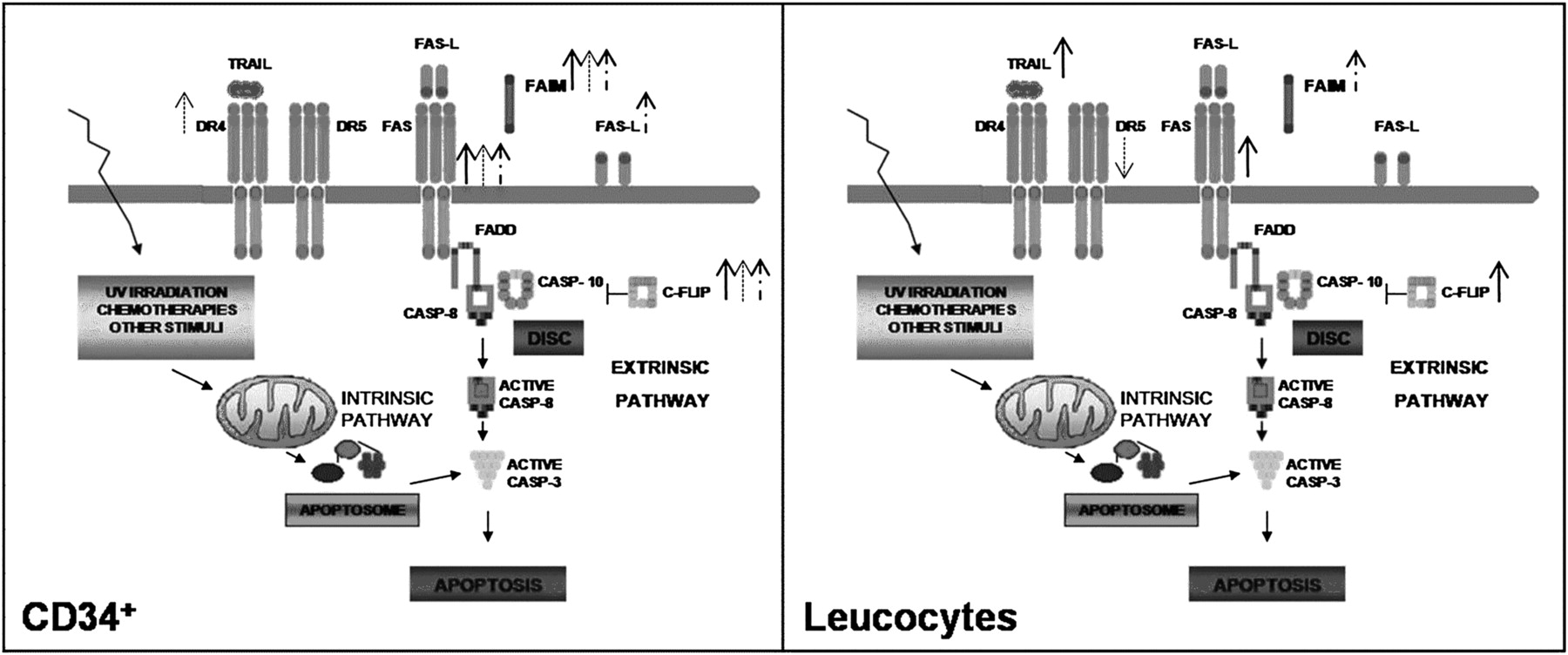{kind=link}
Adaptation of extrinsic apoptosis pathway showing alterations in gene expression in CD34 cells and leukocytes from polycythaemia vera (PV; filled arrows), essential thrombocythaemia (ET; cross-hatched arrows) and primary myelofibrosis (PMF; discontinuous arrows). CASP, caspase.
The detection and monitoring of apoptosis-related gene expression as a biomarker may help in future identification of attractive candidates for new targeted monotherapies or combined therapies for MPD treatment.
Take-home messages
In myeloproliferative diseases (MPDs) there is deregulation of expression of apoptosis-related genes from the extrinsic pathway (fas, fasl, faim, c-flip, trail, dr4 and dr5).
Deregulation of expression of apoptosis-related genes from the extrinsic pathway may contribute to the MPD pathogenesis, leading to cell apoptosis resistance.
fasl and dr5 expression correlate with JAK2V617F mutation in patients with polycythaemia vera.
Acknowledgments
We thank Marcella Grando and Zita Gregório for their technical assistance and help in collecting peripheral blood samples. We are also grateful to Amélia Regina de Albuquerque for help with creating the figures, and Fabiana Rosseto de Morais and Patrícia V. B. Palma for assistance with flow cytometry assays.
References
Supplementary materials
Web Only Data JCP.2010.080895
Footnotes
Funding RT is the recipient of a fellowship from the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (No 08/54387-5), EPLG and RPN are recipients of fellowships from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), and GLVO is the recipient of a fellowship from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). This work was supported by a FAPESP grant (No 06/50094-8).
Ethics approval This study was conducted with the approval of the Brigadeiro Hospital and Hospital das Clínicas of the University of São Paulo at Ribeirão Preto School of Medicine ethics committees (No 014/06 and No 10529/2006, respectively).
Provenance and peer review Not commissioned; externally peer reviewed.