Article Text

Abstract
Background/Aims—The pathogenesis of idiopathic pulmonary fibrosis (IPF)/usual interstitial pneumonia (UIP), a chronic and incurable human respiratory disease, is not well established. This study was designed to investigate whether the apoptosis of type II pneumocytes could be the precipitating factor in the pathogenesis of IPF.
Methods—Nineteen specimens obtained by retrospective review of the medical and pathological records of 55 patients with IPF, four normal subjects, and 10 disease control lungs were analysed. The selected specimens had normal alveoli with intervening patchy scarring of the lung parenchyma, fulfilling the pathological criteria for UIP. To identify individual cells undergoing apoptosis in the normal alveoli, electron microscopy and in situ end labelling of fragmented DNA were performed on paraffin wax embedded sections using digoxigenin-11-dUTP and the enzyme terminal deoxynucleotidyl transferase.
Results—Apoptosis was detected in the normal alveoli of 17 of the 19 patients with IPF/UIP and was absent in the controls. Electron microscopy demonstrated apoptotic changes in type II pneumocytes. These results indicate that apoptotic type II pneumocyte death occurs in normal alveoli of IPF/UIP and could be the principal cause of several events that account for the histological, clinical, and functional alterations seen in IPF/UIP.
Conclusions—In conclusion, numerous type II pneumocytes from the normal alveoli of most patients with IPF/UIP actively undergo programmed cell death. This finding may shed new light on the pathogenesis of this disease, with implications mainly for the treatment of affected patients.
- apoptosis
- type II pneumocytes
- idiopathic pulmonary fibrosis
- pathogenesis
Statistics from Altmetric.com
Programmed cell death, or apoptosis, is a highly regulated and active process that contributes to the control of cell number during development and to the maintenance of many adult tissues.1–3 It is triggered by the activation of an internally encoded suicide programme as a result of either extrinsic or intrinsic signals.4 Morphologically, apoptosis is different from necrosis: it is characterised by blebbing of the cell membrane, a reduction in cell volume, condensation of nuclear chromatin, and endonucleolytic degradation of DNA at nucleosomal intervals.2 Apoptotic bodies are digested or phagocytosed by adjacent cells or macrophages without inducing an inflammatory response.
The crucial role of apoptosis in pathological conditions is increasingly being recognised.5, 6 Recently, components of the Fas–Fas ligand immunological pathway have been detected in patients with idiopathic pulmonary fibrosis (IPF)/usual interstitial pneumonia (UIP).7 IPF/UIP is an idiopathic condition that leads to a progressive reduction in lung volume, shortness of breath, restrictive lung function, and impairment of carbon monoxide diffusing capacity/gas transfer. The pathogenesis of IPF has been the subject of extensive investigation. Several studies have suggested that IPF begins as an alveolitis, which is followed by the development of intraluminal fibrosis and incorporation into the interstitium, as the result of fibroblastic proliferation and collagen deposition.8–11 Ultrastructural studies in patients with the acute form of interstitial pneumonia have shown epithelial necrosis, followed by collapse and apposition of the alveolar walls, to be an important mechanism of parenchymal remodelling.12 Evidence that the same mechanism also contributes to the pathogenesis of IPF was proposed by Myers and Katzenstein.13
Studies on normal lungs show that 7% of the alveolar surface is covered by type II pneumocytes, which make up 16% of the total alveolar cells. Larger type I pneumocytes are flattened and cover approximately 93% of the epithelial surface, representing 8% of the total cells.14 Type II pneumocytes, but not type I cells, are capable of undergoing mitosis and transformation into type I pneumocytes.15 The synthesis, storage, and secretion of surfactant is the function of type II cells. A complex of phospholipids and proteins, surfactant is a thin film that lines the alveolar surface and drastically decreases surface tension of the air–tissue interface, thereby maintaining alveolar stability.
Apoptosis of type II epithelial cells could interfere with the synthesis of surfactant, thereby allowing alveolar collapse. Type I pneumocytes would also be reduced in number, causing denuding of the alveolar capillary membrane, and leading to the development of fibrosis and parenchyma remodelling.
In this study, we investigate the hypothesis that apoptosis of type II pneumocytes could be the first and leading cause of several events that result in IPF/UIP.
Methods
STUDY POPULATION
The study population was identified by retrospective review of the medical and pathological records of 55 patients with IPF at the “Hospital das Clínicas” from 1985 to 1998. In all subjects, lung tissue was obtained by open lung biopsy from two different sites, most often the upper and lower lobes of the same lung. All subjects were evaluated clinically, radiographically, and by lung function tests. All diagnoses were established according to previously described clinical and histological criteria.8, 9, 16, 17
MORPHOLOGICAL STUDY
The tissue specimens were immediately fixed in 10% neutral buffered formalin and embedded in paraffin wax. Six sections (6 μm thick) from each paraffin wax block were stained routinely with haematoxylin and eosin. The slides were examined using light microscopy and classified as UIP according to the previously defined criteria.17 Because our working hypothesis was that apoptosis in the normal alveoli can precede end stage scarring, only the cases with extensive normal alveolar areas were included in our study. Thus, 19 patients (15 men and four women; mean age, 59 years) with IPF, pathologically classified as UIP, were selected because of the presence of normal or nearly normal alveoli with intervening patchy scarring of the lung parenchyma (fig 1A). Patients with connective tissue disease (a common cause of UIP), drug or chemical exposure, or other possible aetiologies of interstitial lung disease, such as bronchiolitis obliterans and diffuse alveolar damage, lymphocytic interstitial pneumonia, bronchiolitis obliterans, and organising pneumonia, or unclassifiable forms of chronic interstitial pneumonia were excluded from our study. Normal control lungs were from four subjects (three men and one women; mean age, 41 years) who were victims of sudden traumatic death (two accidents, one homicide, and one suicide). To ensure that lungs were collected with minimal autolytic changes, we selected victims whose necropsy was performed within the first six hours after death. Ten patients (three women and seven men; mean age, 60 years; SD, 3.6) who had undergone resection for lung cancer were included as disease controls. Tissue specimens were obtained from areas without pneumonia, tumour, or emphysema. None of these control subjects met any of the histology criteria for pulmonary fibrosis.
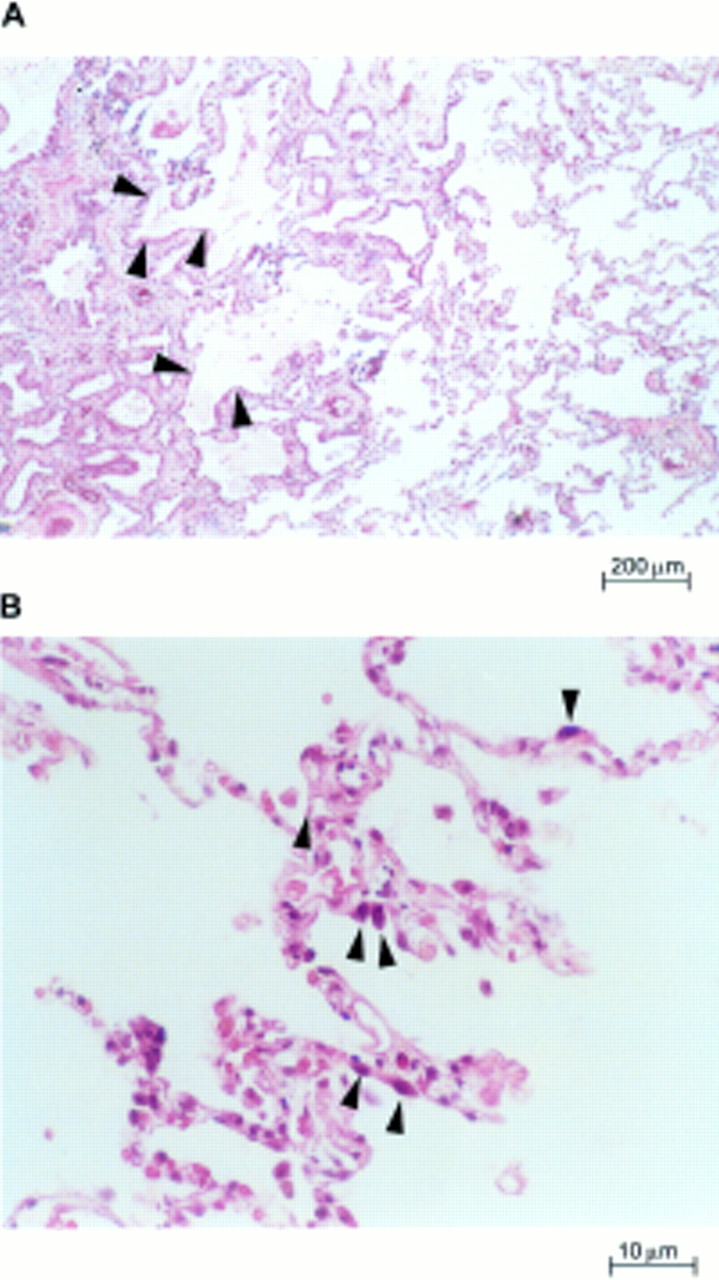
Lung parenchyma of patients with usual interstitial pneumonia. (A) Characteristic variation in histological appearance from one area to another. Dense collagen deposition in the lung parenchyma is seen on the left, and patchy areas containing normal or nearly normal alveoli are present nearby (right and bottom centre). There is a zone of microscopic honeycomb change at the middle left (arrowheads) that is characterised by mucin filled, enlarged air spaces separated by fibrosis (haematoxylin and eosin staining; magnification, ×100). (B) High magnification view of the surviving zones of alveoli showing marginated masses of chromatin within shaped, rounded alveolar cells with eosinophilic cytoplasm (arrowheads), suggestive of type II pneumocytes (haematoxylin and eosin staining; magnification, ×400).
Electron microscopy was performed on samples from two patients (a man and woman; 59 and 65 years old, respectively) who died from pulmonary fibrosis and whose biopsies were included in our study. Tissues were fixed in 2% buffered glutaraldehyde, embedded in Araldite, and thin sections were stained with uranyl acetate and lead citrate.
IN SITU DETECTION AND MEASUREMENT OF APOPTOTIC CELLS
For the in situ detection of apoptosis at the level of a single cell we used the deoxynucleotidyl transferase (TdT) method of end labelling (Boehringer Mannheim, Mannheim, Germany).18, 19 This method involves the addition of deoxyuridine triphosphate (dUTP) labelled with fluorescein to the ends of the DNA fragments by the catalytic action of TdT. All the end labelling experiments were performed several times so that the results for various tissue samples, including rat prostate, could be standardised. Paraffin wax embedded sections (4–6 μm) were layered on to glass slides. The tissue sections were dewaxed with xylene and rehydrated with graded dilutions of ethanol in water. The slides were washed four times with double distilled water for two minutes and immersed in TdT buffer (Boehringer Mannheim). The sections were then covered with TdT (0.3 U/μl) and fluorescein labelled dUTP in TdT buffer, and the samples were incubated in a humid atmosphere at 37°C for 60 minutes. For negative controls, TdT was eliminated from the reaction mixture. The sections were then incubated with antibody specific for fluorescein conjugated to peroxidase. The stains were visualised with a substrate system in which nuclei with DNA fragmentation stained brown. The reaction was terminated by washing the sections twice in phosphate buffered saline. The nuclei without DNA fragmentation stained blue as a result of counterstaining with haematoxylin. Positive controls consisted of rat prostate gland after castration. Two sections from each specimen were examined. Quantitation was performed by a conventional point counting procedure,20 using a reticulated eyepiece (100 points and 50 lines). At a magnification of ×100, 10 non-coincident microscopic fields were studied for each sample to estimate the proportion of the section occupied by apoptotic cells in normal and remodelled tissue; a total of 1000 points, covering an area of 1.0 mm2/section, were counted. The error of this procedure was estimated according to Gundersen and colleagues20 and was always less than 5%. After this procedure, the number of points overlying apoptotic nuclei in positive areas of normal and remodelled lung were counted in 10 fields of ×400 magnification, covering an area of 62 500 μm2/field for each sample. This represented the proportion of positive apoptotic nuclei in areas of normal and remodelled tissue. The extension of areas with apoptotic nuclei (Aapop) and proportion of apoptotic nuclei in positive areas (Papop) were obtained by the following relations:
 where PTapop, PTtiss, and PNapop were the number of points overlying tissue with apoptotic nuclei, total tissue (normal or remodelled), and apoptotic nuclei, respectively (as a percentage).
where PTapop, PTtiss, and PNapop were the number of points overlying tissue with apoptotic nuclei, total tissue (normal or remodelled), and apoptotic nuclei, respectively (as a percentage).
For all morphometric procedures, an estimate of the error (CE) was computed by the following relations: where SE represents the standard error of the mean. Because of the number of points counted, CE was kept under 10% in all morphometric measurements.
where SE represents the standard error of the mean. Because of the number of points counted, CE was kept under 10% in all morphometric measurements.
STATISTICAL ANALYSIS
Comparisons between the extent of apoptosis in normal alveoli and remodelled areas and the percentage of apoptotic type II pneumocyte nuclei in these areas in lung sections from patients with IPF/UIP were made with a paired samples t test using the SPSS program, version 6.0.21
Results
In situ end labelling of fragmented DNA with TdT and digoxigenin-11-dUTP did not reveal apoptosis in sections from the four normal and 10 disease control lungs (fig 2A). In the prostatic epithelium of four day castrated rats, used as positive controls, only the apoptotic cells and the apoptotic bodies within the macrophages at the base of the epithelium reacted positively. Sections of lung from 17 of the 19 patients with UIP showed numerous cells with genomic DNA fragments in their nuclei (fig 2B). Under a light microscope at high magnification most of these cells appeared to be type II pneumocytes because they were well shaped, rounded, and had eosinophilic cytoplasm (fig 1B). Apoptotic type II pneumocytes were frequently found in regions of lung with intervening normal or nearly normal alveoli. They were found in small numbers in remodelled regions replaced by fibrous tissue. Both the extent of the regions with apoptotic cells and the number of apoptotic nuclei varied among the patients (table 1). There was a significant difference (p = 0.0001) between the proportion of regions with apoptotic cells and the number of apoptotic nuclei available from normal regions compared with remodelled regions.
Quantitative analysis data and comparison between the extent of apoptosis in normal alveoli and remodelled areas and the proportion of apoptotic type II pneumocyte nuclei in these areas in sections of lung from patients with IPF/UIP

In situ end labelling of fragmented DNA with terminal deoxynucleotidyl transferase and digoxigenin-11-dUTP. Cells with fragmented DNA stained brown, whereas cells with normal nuclei stained blue (immunoperoxidase staining with haematoxylin counterstaining). (A) A section from a normal lung shows no apoptotic nuclei (magnification, ×160). The intra-alveolar cells seen are macrophages containing carbon pigment (arrowheads). (B) Lung sections from patients with usual interstitial pneumonia (magnification, ×400) with increased labelling. Numerous alveolar nuclei undergoing apoptosis suggestive of type II pneumocytes (arrowheads).
The detection of apoptotic cells by in situ end labelling of fragmented DNA was supported by the fact that pathological criteria for apoptosis were also evident on electron and light microscopy. In sections stained with haematoxylin and eosin, the nuclei of numerous type II pneumocytes showed marginated masses of chromatin, along with discrete, well preserved apoptotic nuclei—typical pathological features of apoptosis (fig 1B). No sign of apoptosis was seen in adjacent sections stained with haematoxylin and eosin from the four normal subjects and the 10 controls with lung disease (data not shown).
ELECTRON MICROSCOPY OF APOPTOTIC TYPE II PNEUMOCYTES
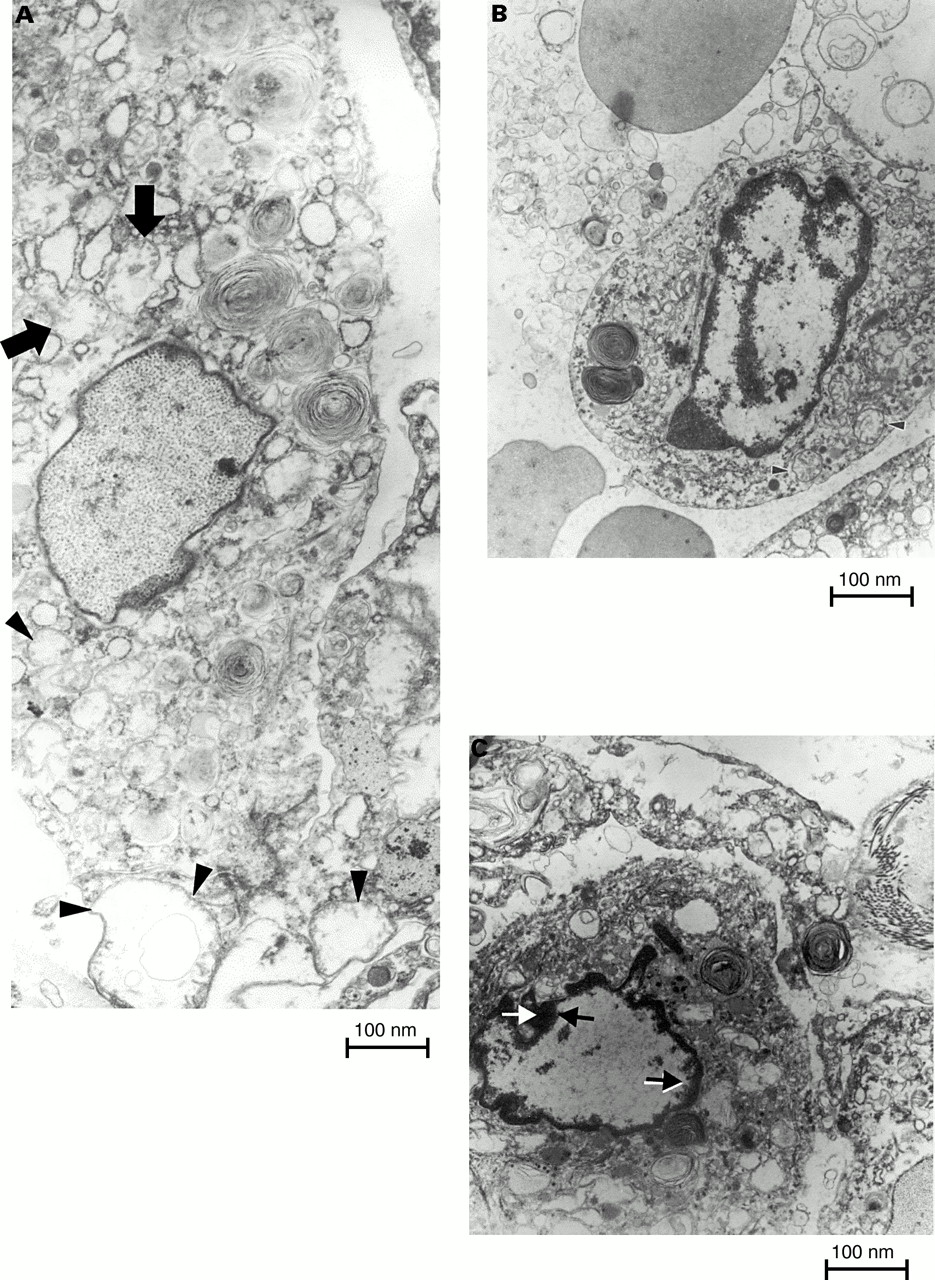
Figure 3A–C shows an electron microscopical view of type II pneumocytes. In non-explicit apoptosis, type II pneumocytes have fine and uniformly dispersed chromatin within the nuclear membrane. These pneumocytes may have swollen mitochondrial profiles and may exhibit rupture of the outer membrane because of the pressure of the inner mitochondrial membrane, which surrounds a greatly swollen inner mitochondrial compartment (fig 3A; arrows and arrow heads). The latter mitochondrial structural alterations are considered to precede those apoptotic stages in which chromatin condenses.22–25 Thus, the cell in fig 3A might be irreversibly committed to die. Figure 3B and C shows type II pneumocytes with very different nuclear profiles to those of non-apoptotic cells. Both figures show progressive condensation of the chromatin, a clarifying process of the remaining nuclear region, and a final degradation of the cytoplasmic structure.

{kind=link}
{kind=link}
{kind=link}
Sections of lung from patients with usual interstitial pneumonia examined by electron microscopy. (A) Type II pneumocytes undergoing non-explicit apoptosis, characterised by fine and uniformly dispersed chromatin within the nuclear membrane. These pneumocytes have swollen mitochondrial profiles, sometimes with rupture of the outer membrane because of the pressure of the inner mitochondrial membrane, which surrounds a greatly swollen inner mitochondrial compartment (arrows and arrows heads). (B and C) Type II pneumocytes with very different nuclear profiles to those of non-apoptotic cells, characterised by progressive condensation of the chromatin as part of a clarifying process of the remaining nuclear region and a final degradation of the cytoplasmic structure (magnification, ×9000).
Discussion
We report the occurrence of apoptotic alveolar cell death in IPF. Uhal and colleagues5 suggested that apoptosis might be a mechanism of cell death in fibrosing lung diseases. Since then, there has been a rapidly growing and relevant literature on apoptosis in acute lung injury and experimental models of lung fibrosis have been developed.7
In our study, apoptosis was identified by in situ end labelling of fragmented DNA with TdT and digoxigenin-11-dUTP, a commonly accepted method for the detection of the apoptotic process.18, 19 There has been concern about the ability of such in situ labelling methods to distinguish between cell necrosis and apoptosis. However, in our study the detection of a positive reaction with in situ end labelling was correlated with the presence of typical signs of apoptosis (masses of chromatin and well preserved apoptotic bodies) in sections stained with haematoxylin and eosin.19 Moreover, the absence of an inflammatory reaction in regions of lung with intervening normal or nearly normal alveoli in most of our patients argues against necrosis and for apoptosis. Furthermore, no positive staining was detected in alveolar cells from age matched normal subjects whose lungs were processed in the same manner as those of our patients, and the in situ end labelling of fragmented DNA was not detected when the enzyme TdT was omitted.
We found that numerous cells in the normal alveoli of patients with idiopathic pulmonary fibrosis underwent apoptosis (fig 1B and fig 2B). Most of these cells were morphologically similar to type II pneumocyte cells under high power magnification. Further evidence of apoptotic changes in type II pneumocytes was gained by electron microscopy (fig 3A–C). Clumping of chromatin at the inner nuclear membrane is a hallmark of apoptosis. The evaluation of nuclear alterations by electron microscopy is thought to be the best way of detecting programmed cell death.1 An early stage of explicit apoptosis is the disassembly of the nuclear lamina, a scaffold where the euchromatin loops anchor. Consequently, these loops collapse and there is a progressive increase in the amount of condensed perinuclear chromatin and a corresponding decrease in the density of euchromatin normally represented by a delicate texture of granule filamentous material. This clarification of a sector of the apoptotic nuclei progresses to the point where this region looses most of its electron density, eventually becoming translucent. These changes can be seen in many of the figures of Kerr and colleagues,1 and also in other electron microscopic images of advanced, non-fragmented, apoptotic nuclei. One of the reasons that not all apoptotic nuclei have a transluscent euchromatic region is that during explicit apoptosis the nucleus tends to shrink, thereby concentrating the contents of its various parts. However, as far as we know, clarification of the nuclear content (as seen in the dead cell in fig 3C) has not been ascribed to necrosis.1
The remodelled areas had few or no apoptotic cells, whereas apoptotic nuclei were frequently seen in areas with little involvement. This finding suggests that the loss of type II pneumocytes through apoptosis is a primary process that precedes the filling of acellular space by fibrous tissue in the absence of an inflammatory reaction.
The extent and the number of apoptotic type II pneumocytes varied among the patients (table 1). The absence of apoptosis in two of our patients is intriguing. The clinical and histological features of these patients were similar to those of the others, but perhaps their disease was in remission.
The triggering factors for apoptotic type II pneumocytes in IPF/UIP remain to be elucidated. Fas ligand was detected in inflammatory cells in bronchoalveolar lavage fluid (BALF) from patients with IPF.7 In addition, the expression of Fas appeared to be upregulated in bronchiolar and alveolar cells. However, in that study there was no indication of the alveolar epithelial cell type expressing Fas. This mechanism appears not to be involved in our cases because the alveoli with apoptotic cells do not contain inflammatory cells.
We predicted that excessive apoptosis might cause damage to type II pneumocytes, as deduced from nuclear changes and signs of cytoplasmic degeneration (figs 2B and 3A–C). This finding leads us to hypothesise that type II cell apoptosis plays an important pathological role as an early lesion in IPF, raising important questions about the mechanisms involved in the pathogenesis and clinical correlation of the disease.
First of all, the impact of apoptosis on the surfactant system could have several consequences on alveolar histoarchitecture. These apoptotic features could be the morphological expression of impaired release and/or defective surfactant production, with quantitative and qualitative alterations in the surfactant system. This abnormal or absent surfactant leads to alveolar collapse, alveolar wall apposition, and compensatory alveolar duct enlargement—the alveolar ducts do not collapse because of the interdependence between adjacent units.26 The surfactant system abnormalities that occur with IPF have strong support from the literature. Robinson and co-workers27 studied 15 untreated patients with IPF diagnosed by open lung biopsy and found less than half the amount of phospholipid in BALF from these patients compared with healthy volunteers. In addition, patients with IPF had a lower proportion of phosphatidyl glycerol and a higher proportion of phosphatidyl inositol in the BALF than did healthy volunteers. The authors correlated pronounced changes in phospholipid composition with more advanced fibrotic processes in the histopathology of the disease. In another report, Mc Cormac and co-workers28 measured surfactant protein A (SP-A) in BALF from 44 patients with IPF and 33 healthy volunteers. The mean SP-A/total phospholipids (PL) was lower in patients with IPF than in healthy volunteers. They concluded that SP-A/PL is a biochemical marker in BALF that might predict survival in patients with IPF/UIP.
The implications of alveolar collapse in the pathogenesis of IPF/UIP have been emphasised by Burkhardt,29 Katzenstein,12 and Myers et al.13 In addition, alveolar collapse, more so than fibrosis, explains the findings of Coxson and et al.30 These authors quantified lung structure in IPF by computerised tomography and histological analysis. The results showed that in contrast to control lungs IPF was associated with a reduction in airspace volume, without changes in tissue volume or weight. Lung surface area decreased two thirds and mean parenchymal thickness increased 10-fold. They concluded that the architectural rearrangement of lung tissue in IPF lungs contributed to the loss of airspace and surface area without increasing total lung tissue. Reinforcing this concept, the collagen content and morphological degree of fibrosis were quantified in nine patients with IPF and in six control subjects by Fulmer and et al.31 They found no significant difference in collagen content among IPF and control lungs. Equally, there was no correlation between the morphological assessment of the degree of fibrosis and collagen content. The results of this study are consistent with the concept that IPF is a disease involving alterations in quality, form, and location of collagen, rather than simply a disease of increased interstitial collagen. Furthermore, recent evidence has shown that the absence of prostaglandin E2,32, 33 usually produced by type II alveolar cells, enhances fibroblast growth and collagen synthesis.34 Nevertheless, delayed repair of the alveolar epithelium disturbs normal epithelial–fibroblast interactions and is sufficient to promote the fibroblastic process.35 The “fibroblast foci” described by Katzenstein and Myers36 must correspond to the fibrosis, which appears in the denuded areas of alveolar membrane resulting from type II pneumocyte apoptosis. Another consequence of type II pneumocyte apoptosis might be related to the turnover of type I pneumocytes. Because type I pneumocytes are produced by the proliferation and transformation of type II pneumocytes, the progressive depletion of the latter leads to a proportional reduction in the number of type I pneumocytes.
The missing type I pneumocytes in the alveolar duct and respiratory bronchiole are replaced by the proliferation of cuboidal epithelial cells derived from bronchiole basal cells (type A cuboidal cells) and from cuboidal cells from the respiratory bronchiole (type B cuboidal cells),37 resulting in a honeycomb configuration. Thus, if type II pneumocyte apoptosis is the pathogenic mechanism involved in IPF, it is difficult to understand the hypertrophy and hyperplasia of these cells to renew the alveolar surface, as reported by Kawanami and et al.37 On the other hand, it is currently accepted that type II pneumocyte proliferation and transformation occur after type I pneumocyte necrosis,15 as in acute interstitial pneumonia. In the latter, the histological hallmark is interstitial fibrosis and oedema associated with type II pneumocyte hyperplasia.38 Furthermore, in non-specific interstitial pneumonia/fibrosis type II pneumocyte hyperplasia occurs in 50% and 44% of the cases, respectively.39 It may be that the patients in Kawanami's report37 with hypertrophy and hyperplasia of type II pneumocytes in “areas of less severe degree of fibrosis” have non-specific interstitial pneumonia/fibrosis, as recently described by Katzenstein and colleagues,39 and now included in the new IPF classification.36 Thus, based on the above considerations, we think that the presence of type II pneumocyte hyperplasia in lung biopsy specimens exhibiting an interstitial process should suggest a diagnosis other than UIP.
Because type II pneumocyte apoptosis might be involved in IPF/UIP pathogenesis, it is reasonable to suppose that the presence of alveolitis8–10 is not a necessary prelude to IPF. As previously emphasised by Carrington et al,17 we must be careful in accepting the concept proposed by Tubbs et al,40 who described desquamative interstitial pneumonia (DIP) as the initial or cellular phase of IPF. This finding has strong support in the recent report of Hartman et al.41 They studied radiological progression in treated patients with UIP and DIP and found that these two entities have different radiological behaviour. According to Katzenstein et al,36 in UIP “the inflammation occurs mainly in areas of collagen deposition or honeycomb change, and rarely in otherwise unaltered alveolar septa. There is no evidence that inflammation is more prominent in early disease. The presence of severe inflammation, in fact, should suggest a diagnosis other than UIP.”.
Finally, we must emphasise that UIP progresses by recurrent episodes of type II apoptosis in new pulmonary parenchymal areas. This observation is derived from the concomitant presence of fibrotic areas and normal alveoli where the process starts by type II pneumocyte apoptosis.
In conclusion, we found that numerous type II pneumocytes from the normal alveoli of most patients with IPF/UIP actively undergo programmed cell death. This finding sheds new light on the pathogenesis of this disease, which might have implications for patient treatment. In the future, steroid treatment, which is successful in less than 20% of patients with IPF/UIP,42 should be replaced by substances with inhibitory action in type II pneumocyte apoptosis.
Acknowledgments
We are indebted to the scientific department of FMUSP for the helpful preparation of the photomicrographs. This work was supported by the Brazilian funding agencies: FAPESP, CAPES, CNPq, and LIM-HCFMUSP.