Article Text

Abstract
Background/Aims—Fas ligand (FasL) is a mediator of apoptosis via the Fas receptor (Fas/CD95/APO-1). Normal colonic epithelium expresses Fas, and appears to be relatively sensitive to Fas mediated apoptosis. Colonic adenocarcinomas coexpress FasL and Fas without undergoing widespread apoptosis. This study investigates the expression of FasL in colonic carcinogenesis from the earliest stages of the adenoma–carcinoma sequence.
Methods—FasL expression was determined in colonic adenomas (n = 38) of varying degrees of dysplasia and histological type by immunohistochemistry. Adenomas that contained areas of carcinomatous change were included (n = 12 of 38). Normal colonic epithelium (n = 10), hyperplastic polyps (n = 8), and serrated adenomas (n = 3) from patients without colonic adenocarcinomas were used for comparison. Cell death was detected in situ in adenomas using TUNEL (terminal transferase mediated dUTP nick end labelling).
Results—In normal colonic epithelium and hyperplastic polyps, FasL expression was restricted to the luminal surface of the crypts, where Fas–FasL coexpression was coincident with a high frequency of TUNEL positive epithelial cells. All adenomas (n = 38) had an altered distribution of positive FasL staining; FasL expression was found in most cells (> 70% of neoplastic cells). Expression of Fas was also detected throughout the adenomas, but coexpression of FasL and Fas was not associated with TUNEL positivity in most cells.
Conclusions—FasL upregulation occurs early in the adenoma–carcinoma sequence of colon carcinogenesis, and is evident at the level of mild dysplasia. The lack of pronounced apoptosis in areas of adenomas coexpressing Fas and FasL suggests that colonocytes acquire resistance to Fas mediated apoptosis early in the transformation process.
- Fas (CD95/APO-1)
- Fas ligand
- colon
- adenoma
Statistics from Altmetric.com
Most colonic adenocarcinomas develop from adenomatous polyps. Colonic adenomas are foci of dysplastic epithelium (intraepithelial neoplasia) and approximately 5% of colonic adenomas become malignant. These adenomas become gradually more dysplastic over a period of several years and ultimately develop into cancer.1–3 This colonic adenoma–carcinoma sequence is a relatively well defined example of multistep carcinogenesis. As genetic alterations accumulate in the transition from normal tissue to neoplasia, the tissue phenotype alters from normal to adenomatous polyps with varying degrees of dysplasia.4 In contrast to adenomatous polyps, hyperplastic polyps are composed of well defined glands and crypts lined by non-neoplastic columnar epithelium and have no well defined premalignant potential. Serrated adenomas are benign neoplastic lesions with mixed histological features of both hyperplastic and adenomatous polyps. Serrated adenomas have some malignant potential and are managed clinically like adenomatous polyps.5, 6
The Fas (CD95/APO-1) receptor (Fas) is a cell surface receptor that mediates apoptotic cell death upon triggering by Fas ligand (FasL). FasL expression was initially thought to be restricted to various lymphoid cells, including T cells, B cells, natural killer cells, and neutrophils. Recently, the expression of FasL has also been reported in several non-lymphoid cells.7–12 Fas is widely expressed in most cell types, including epithelial cells, and Fas mediated apoptosis may be involved in cell turnover in many tissues.13–17 FasL also contributes to immune privilege in certain tissues where it is expressed, by triggering the apoptosis of infiltrating inflammatory cells.18, 19
Fas is expressed in normal colonic epithelium, and normal colonic epithelial cells are relatively sensitive to Fas mediated apoptosis.20 Whereas Fas is downregulated in a proportion of colonic adenocarcinomas,21 FasL is upregulated in most colon tumours.22 Coexpression of Fas and FasL occurs throughout large areas of colon tumours, yet this does not lead to increased apoptosis of the tumour cells. This is in agreement with the finding that Fas expressed in colon carcinoma cell lines fails to transduce an apoptotic signal upon engagement with FasL.23
The aim of our study was to determine at what stage of the adenoma–carcinoma sequence the upregulation of FasL occurs. We examined the expression of FasL by epithelial cells in normal colonic mucosa, hyperplastic polyps, and premalignant adenomatous polyps of varying degrees of dysplasia through the development of carcinoma. We sought to determine whether FasL upregulation was associated with epithelial cell apoptosis.
Materials and methods
TISSUES
Normal human tissues were collected after investigative endoscopic biopsy (n = 10) or resection of colonic carcinomas (n = 10) at the Mercy Hospital, Cork. Normal specimens from patients undergoing surgical resection of colonic carcinoma were taken from the disease free resection margins and on routine haematoxylin and eosin staining were entirely normal morphologically, with no metaplasia or dysplasia evident. Serrated adenomas (n = 3) (table 1), hyperplastic polyps (n = 8) (table 2), and colonic adenomas (n = 38) (table 3) were collected after investigative endoscopic biopsy. Areas of peritumoral epithelium were examined from resected colonic carcinomas not arising in adenomas (n = 32). All specimens were collected according to a protocol approved by the ethics committee of the Cork University teaching hospitals.
Patient population
Size of metaplastic polyps
Adenomas: histological size and type
immunohistochemical detection of fasl and fas
Paraffin wax embedded sections of normal colon and colonic adenomas were dewaxed in xylene and rehydrated before analysis. Slides were washed twice for five minutes in a wash buffer containing 50mM Tris/HCl (pH 7.6), 50mM NaCl, and 0.001% saponin, and endogenous peroxidase was quenched with 3.0% hydrogen peroxide in methanol for five minutes. Slides were washed as before except that the wash buffer for this and all subsequent steps included 1% normal goat serum. Slides were then blocked for one hour in wash buffer containing 5% normal goat serum. Slides were washed and incubated overnight at 4°C with affinity purified, rabbit polyclonal antihuman FasL or Fas specific IgG (Santa Cruz Biotechnology, Santa Cruz, California, USA) at 0.1 g/ml in wash buffer. Antibody binding was localised using a biotinylated secondary antibody, avidin conjugated horseradish peroxidase (HRP), and diaminobenzidine (DAB) substrate, contained within the Vectastain ABC detection kit (Vector Laboratories, Burlingame, California, USA). The appropriate immunising peptide to which the antibody was raised (FasL, amino acids 260–279; Fas, amino acids 316–335; Santa Cruz Biotechnology) was included at 1 g/ml during primary antibody incubation as a direct, internal competitive control for antibody specificity. In a separate control for specificity of the Santa Cruz FasL specific polyclonal antibody, we found previously that specific antisense oligonucleotide mediated inhibition of FasL expression23 converted SW620 cells from positive to negative by immunohistochemistry with this antibody. The FasL specificity of the Santa Cruz antibody has also been verified by others: it stained FasL mRNA positive but not FasL mRNA negative cell lines,24 and produced a similar staining pattern to other FasL specific monoclonal25 and polyclonal26 antibodies. FasL detection was confirmed using immunohistochemistry with a FasL specific monoclonal antibody (clone G247-4; Pharmingen, San Diego, California, USA), as described above. The monoclonal antibody was used at a concentration of 5 g/ml, and an isotype matched control antibody was also used. Slides were counterstained with haematoxylin.
immunophosphatase detection with fasl specific monoclonal antibody
After dewaxing and rehydration as above, sections were pretreated by microwave irradiation in 0.01M citrate buffer for five minutes at 370 W. The sections were cooled rapidly by immersing in 0.1M phophate buffered saline (PBS). The slides were then incubated with a mouse antihuman monoclonal FasL specific monoclonal antibody (clone G247-4; Pharmingen) at a concentration of 5 g/ml for one hour. The slides were then washed for five minutes, using 0.1M Tris buffered saline (TBS), which was used for all subsequent washes. After incubation with a secondary rabbit antimouse immunoglobulin (Dako Corp, Ely, Cambridgeshire, UK) at a dilution of 1/25 for 30 minutes, washing was repeated. APAAP (alkaline phosphatase conjugated anti-alkaline phosphatase) complex (DO651; Dako Corp), at a dilution of 1/50, was added to the slides for 30 minutes, after which they were washed again. To enhance staining, the secondary antibody incubation and APAAP complex incubation were repeated once as above, but the duration of the incubation steps was reduced to 10 minutes each. Alkaline phosphatase substrate solution was then added to the slides for 10 minutes. The phosphatase substrate used was Fast Red (Sigma Chemical Co, St Louis, Missouri, USA). All incubations were carried out at room temperature. Slides were counterstained with haematoxylin and viewed by light microscopy.
TUNEL IN SITU CELL DEATH DETECTION
Cell death was detected in situ in resected tissues by enzymatic labelling of DNA strand breaks using terminal deoxynucleotidyl transferase (TdT) mediated dUTP nick end labelling (TUNEL; Boehringer Mannheim, GmbH, Mannheim, Germany), according to the manufacturer's instructions. Paraffin wax embedded sections were dewaxed in xylene and rehydrated before analysis. After pretreatment by microwave irradiation in 0.01M citrate buffer for five minutes at 370 W, sections were washed in PBS and endogenous peroxidase activity was blocked by 3% hydrogen peroxide in methanol for 30 minutes. The slides were then washed in PBS, which was used for all subsequent washes. The sections were treated with permeabilisation solution (0.1% Triton X-100 in 0.1% sodium citrate) for 15 minutes. After washing, the labelling reaction was performed using a solution containing TdT, its buffer, and fluorescein dUTP. During this step, slides were coverslipped and incubated at 37°C for 60 minutes in a humidity chamber. TdT was omitted from negative control slides, which were included in each run. To localise cells containing labelled DNA strand breaks, sections were washed and incubated with a sheep antifluorescein antibody Fab fragment conjugated with HRP at 37°C in a humidity chamber for 30 minutes. After washing, colour development was performed by incubating sections with a DAB substrate solution for 15 minutes. After a final wash, sections were viewed by light microscopy. Because the TUNEL technique can falsely label DNA strand breaks in proliferating or necrotic cells, only those cells that were both morphologically apoptotic and TUNEL positive were considered to be undergoing apoptosis.27
Results
fasl is expressed by colonic adenomas
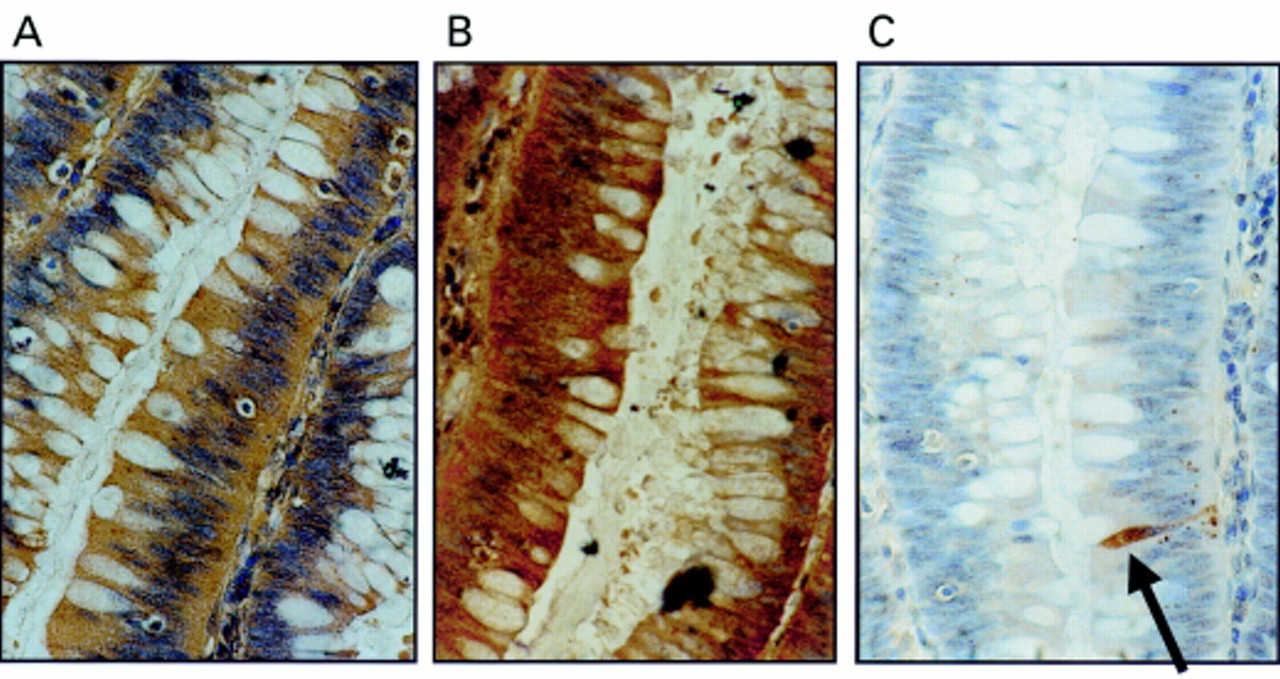
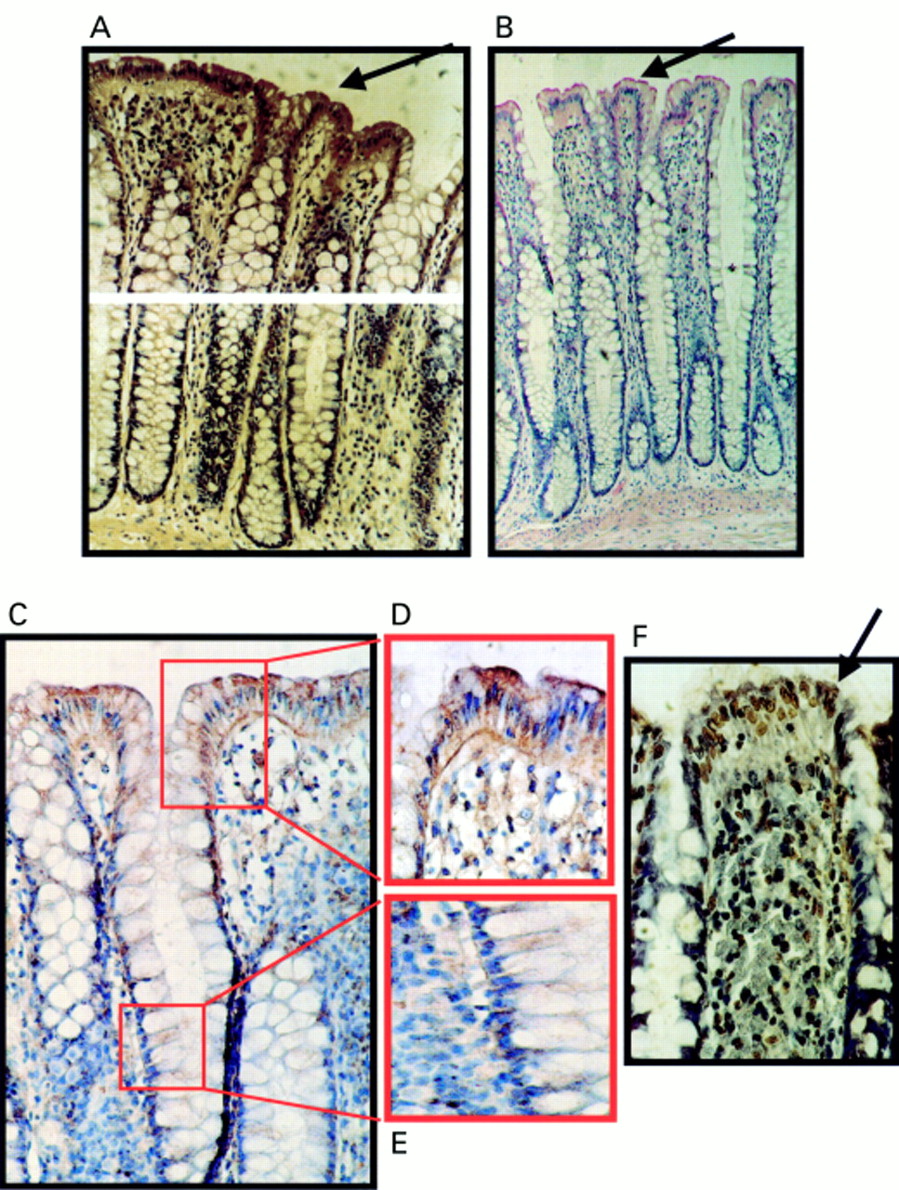
All colonic adenomas examined expressed FasL (fig 1A), irrespective of the degree of dysplasia, adenoma size, or the histological type. FasL was expressed throughout more than 70% of neoplastic cells in all adenomas, and Fas was also expressed by all the adenomas examined (fig 1B). The intensity and extent of Fas staining was more uniform than that of FasL throughout the adenomas. By staining consecutive sections, FasL and Fas were seen to be coexpressed in large areas of most adenomas. Carcinomas arising within adenomas all showed a degree of Fas and FasL staining equivalent to the surrounding adenomas (n = 12 of 38).

Coexpression of Fas ligand (FasL) and Fas in colonic adenomas is not associated with Fas mediated apoptosis. (A) Immunoperoxidase staining using a FasL specific rabbit polyclonal antibody was performed on paraffin wax embedded colonic adenoma sections. Slides were counterstained with haematoxylin. FasL positive immunohistochemical staining (brown) is shown in a representative mildly dysplastic villous adenoma. As a control for specificity of antibody detection, the FasL immunising peptide was included during primary antibody incubation. Competitive displacement of staining by the immunising peptide confirmed FasL specificity (not shown). (B) Another section sequential to that shown in (A) was used to detect Fas, using immunoperoxidase staining with a Fas specific rabbit polyclonal antibody. Fas positive immunohistochemical staining (brown) was present in the adenoma cells. (C) Cell death detection in situ by terminal transferase mediated dUTP nick end labelling (TUNEL). Sections sequential to those shown in (A) and (B) were used to detect cell death by enzymic labelling of DNA strand breaks using TUNEL. Only those cells with positive TUNEL staining (brown) and exhibiting apoptotic morphology (arrow) were considered to be apoptotic. Only occasional TUNEL positive cells were identified despite widespread Fas–FasL coexpression. Control sections (not shown) where the terminal transferase was omitted were negative. These results are representative of 38 colonic adenomas.
Using the TUNEL in situ cell death detection assay, positive cells were distributed throughout the adenomas. However, most cells in areas of adenomas coexpressing FasL and Fas were negative for apoptosis (fig 1C). Hence, coexpression of FasL and Fas failed to result in pronounced Fas mediated autocrine cell suicide or paracrine/juxtacrine cell death. In addition, peritumoral epithelium from 30 of the 32 colonic carcinomas examined showed increased FasL expression (fig 2). In these areas, FasL expression extended into the base of the crypts, in contrast to normal colonic epithelium, where FasL expression was restricted to the luminal surface. Most cells in these areas were non-apoptotic morphologically and by TUNEL.
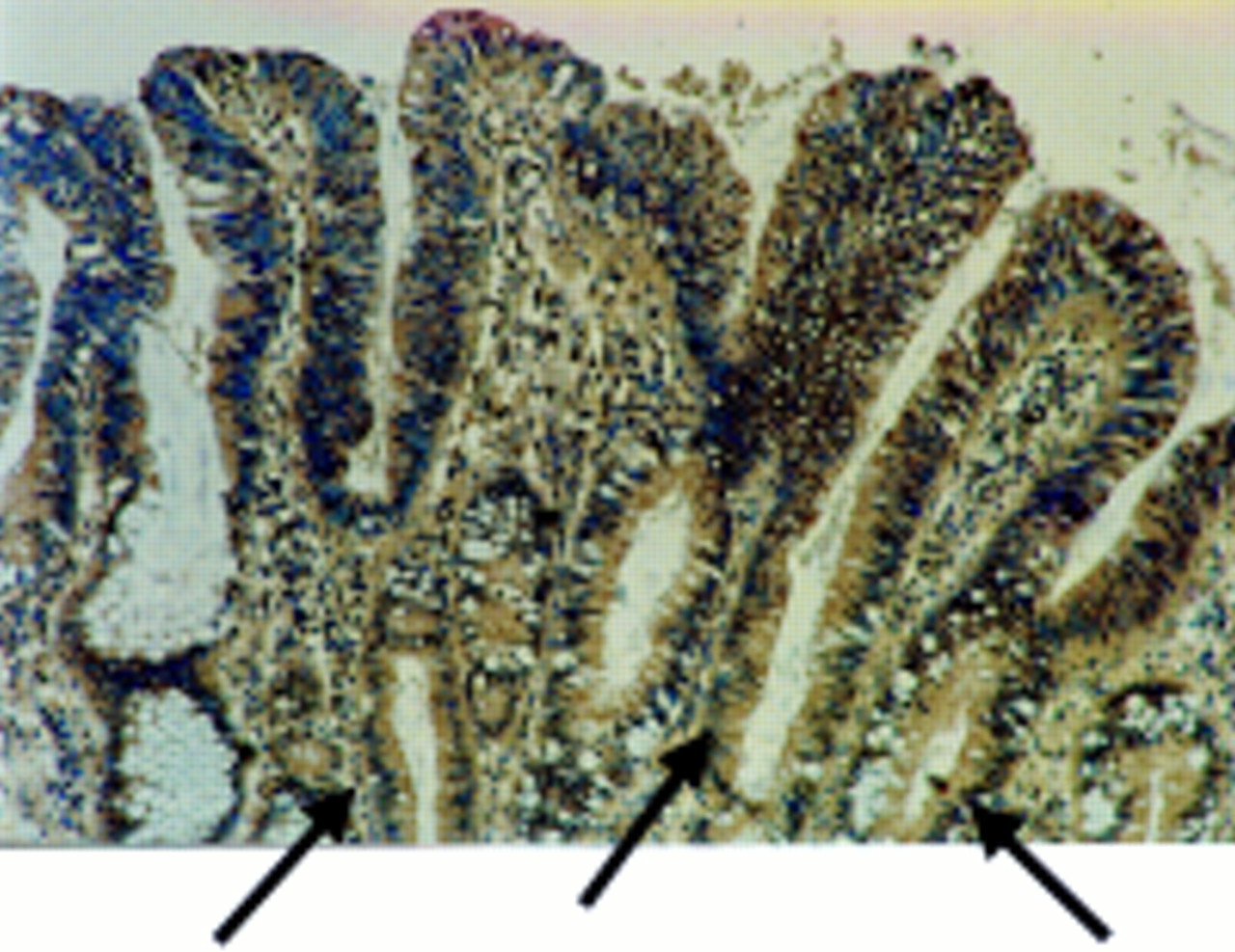
Fas ligand (FasL) is expressed in areas of peritumoral epithelium in colonic adenocarcinomas. Immunoperoxidase staining using a FasL specific rabbit polyclonal antibody was performed on paraffin wax embedded colonic adenocarcinoma sections. Slides were counterstained with haematoxylin. FasL positive immunohistochemical staining (brown) is shown in a representative area of peritumoral epithelium. As a control for specificity of antibody detection, the FasL immunising peptide was included during primary antibody incubation. Competitive displacement of staining by the immunising peptide confirmed FasL specificity (not shown). FasL expression by the neoplastic cells extends into the crypts of the dysplastic glands (arrows). This result is representative of peritumoral epithelium from 30 of 32 colonic carcinomas that showed increased FasL expression.
normal colonic epithelium and hyperplastic polyps express fasl at the luminal surface
A distinct pattern of FasL expression was detected in normal colon epithelium. All normal colonic specimens immunohistochemically stained for FasL were positive (n = 20) (fig 3A–E). The FasL specificity of the polyclonal antibody used (Santa Cruz Biotechnology) was confirmed—staining was inhibited by the inclusion of the immunising peptide as a competitive inhibitor before incubation with the primary antibody. Positive staining of epithelium in normal colonic specimens was restricted to the luminal surface epithelial cells. The maximal intensity of epithelial staining was equivalent to that of FasL positive lymphoid cells observed in all specimens. Crypt epithelial cells were negative for FasL. To confirm the expression of FasL by normal surface colonic epithelial cells, staining was also performed with a different FasL specific antibody (monoclonal antibody clone 247-4; Pharmingen). Results using this FasL specific monoclonal antibody were similar to those obtained with the polyclonal antibody. In addition, FasL staining was independent of secondary detection systems, because similar results were found using either APAAP or immunoperoxidase assays.
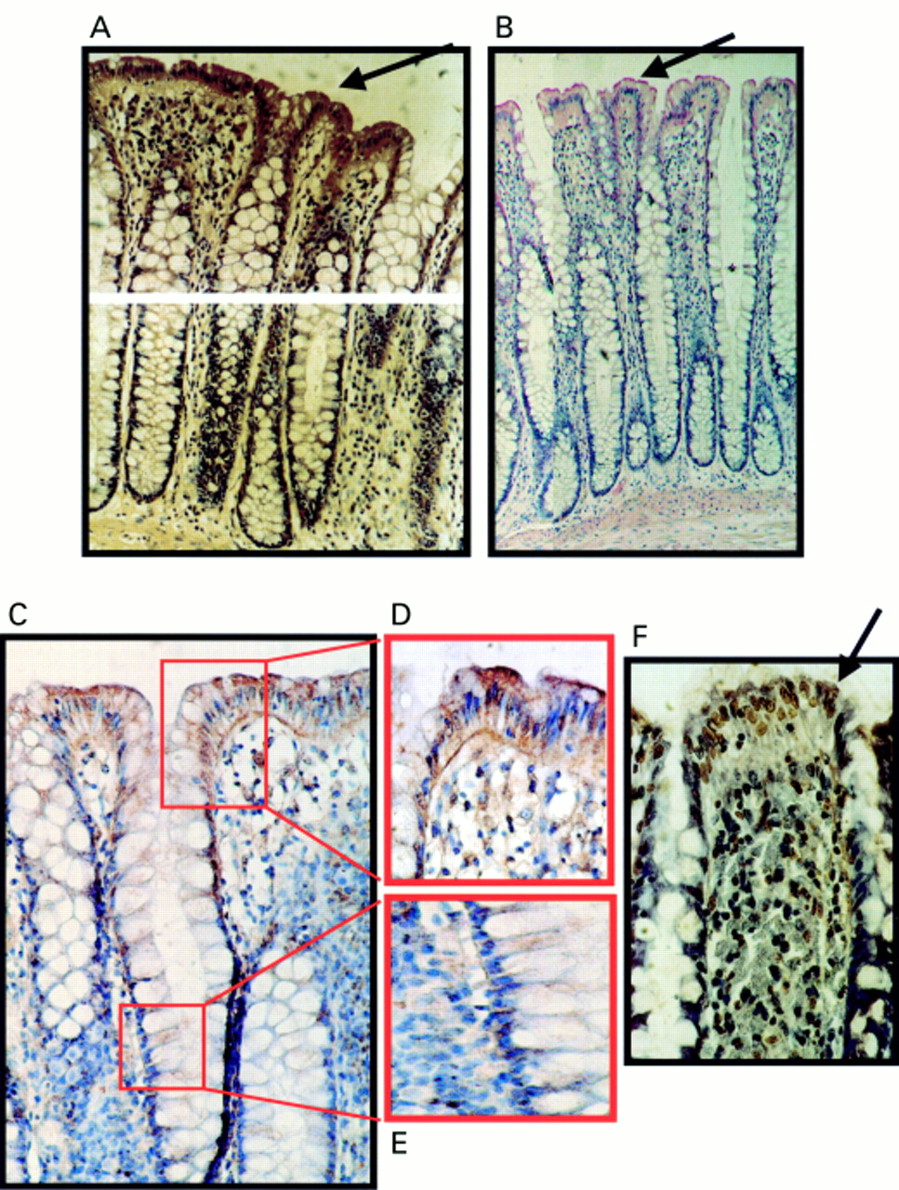
Expression of Fas ligand (FasL) at the luminal surface of normal colonic epithelium coincides with increased frequency of terminal transferase mediated dUTP nick end labelling (TUNEL) positive epithelial cells. (A) Immunoperoxidase staining using a FasL specific rabbit polyclonal antibody was performed on paraffin wax embedded normal colonic sections. Slides were counterstained with haematoxylin. FasL positive immunohistochemical staining (brown) is shown in a representative normal colonic specimen. FasL expression was detected in luminal colonic epithelium (arrow). As a control for specificity of antibody detection, the FasL immunising peptide was included during primary antibody incubation. Competitive displacement of staining by the immunising peptide confirmed FasL specificity (not shown). (B) FasL immunophosphatase (alkaline phosphatase conjugated anti-alkaline phosphatase; APAAP) staining (red) was performed using a FasL specific monoclonal antibody. Slides were counterstained with haematoxylin. Results from staining with the FasL specific monoclonal antibody agreed with those obtained using the polyclonal antibody—FasL positive epithelial cells (red; arrow) were present at the luminal surface. (C) Immunoperoxidase staining with the same FasL specific monoclonal antibody as used in (B) gave identical results. Thus, staining was independent of secondary detection systems because similar results were obtained using either APAAP or immunoperoxidase assays. (D) FasL positive luminal epithelium (brown) from (C) illustrated at higher magnification. (E) Negative crypt epithelium from (C) illustrated at higher magnification. (F) Cell death detection in situ by TUNEL. Cell death was detected by enzymic labelling of DNA strand breaks using TUNEL in normal colonic sections sequential to those shown in (A). Only those cells with positive TUNEL staining (brown) and exhibiting apoptotic morphology (arrow) were considered to be apoptotic. Control sections (not shown) where the terminal transferase was omitted were negative. An increased frequency of TUNEL positive epithelial cells was detected at the luminal surface, where cell death was coincident with FasL expression. These results are representative of 20 normal colonic specimens.
Positive TUNEL staining and characteristic apoptotic morphology confirmed the occurrence of apoptotic cell death in the luminal surface epithelium. TUNEL staining was largely nuclear, as is typical of true apoptotic detection. This suggests that in contrast to adenoma cells, pronounced apoptosis of normal colon epithelial cells coincides with coexpression of Fas and FasL.
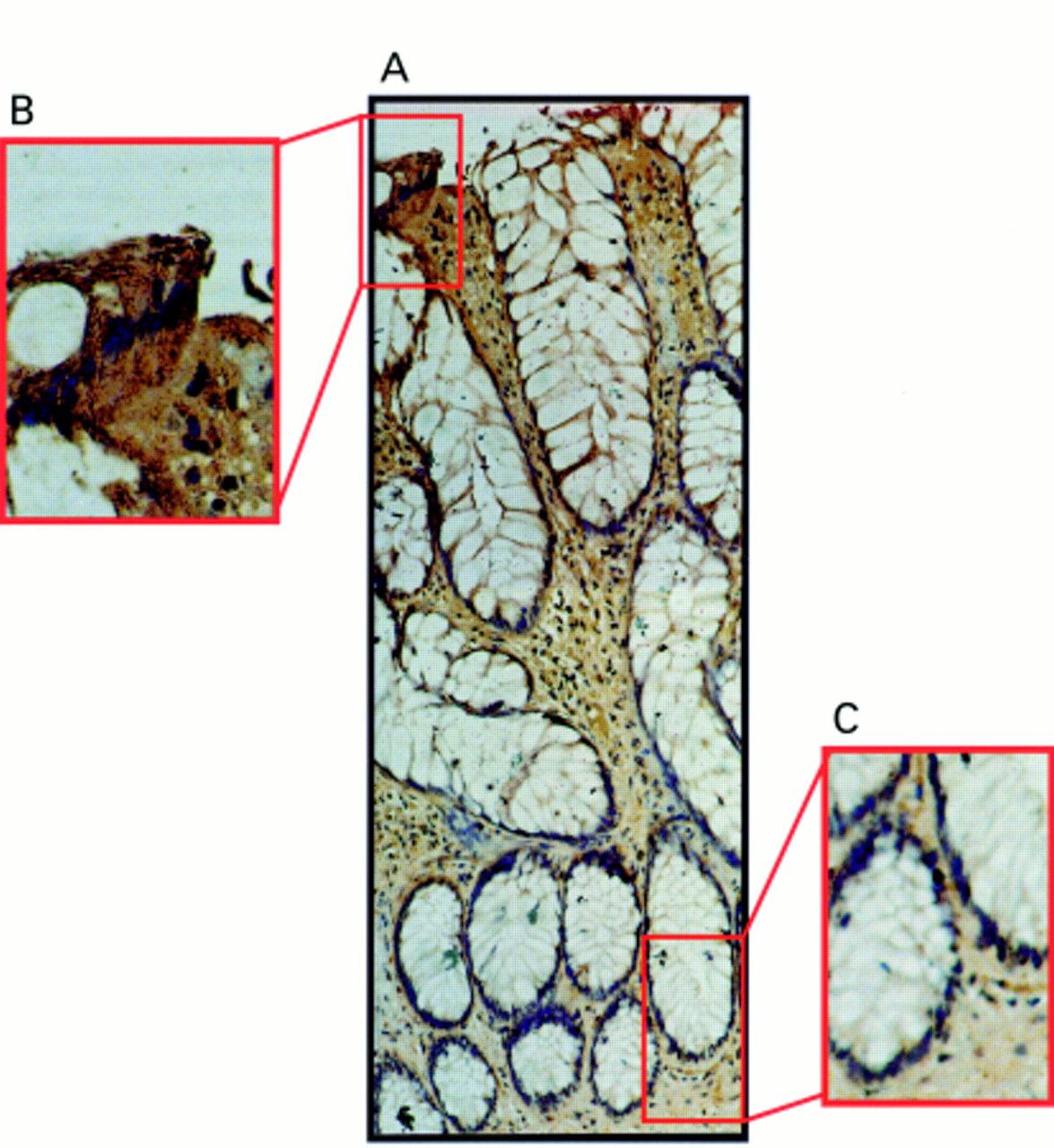
Hyperplastic polyps retained the same pattern of FasL expression as the normal epithelium (n = 8); the luminal epithelium stained positive for FasL whereas crypt epithelial cells were negative (fig 4).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Fas ligand (FasL) expression occurs only at the luminal surface of hyperplastic polyps. (A) Immunoperoxidase staining was performed in paraffin wax embedded hyperplastic polyp sections using a FasL specific polyclonal antibody. Slides were counterstained with haematoxylin. FasL positive immunohistochemical staining (brown) is shown in a representative hyperplastic polyp. The pattern of FasL immunoreactivity was similar to that seen in normal epithelium, with FasL expression occurring in epithelial cells at the luminal surface, but being absent from the crypt base. (B) Detection of FasL positivity in luminal epithelium illustrated at higher magnification. (C) FasL negative crypt epithelium illustrated at higher magnification. These results are representative of 10 hyperplastic polyps.
serrated adenomas showed a mixed pattern of fasl expression
Serrated adenomas showed a mixed pattern of FasL expression, varying locally between the expression pattern of normal colonic epithelium and that of adenomatous polyps. FasL was restricted to the luminal surface throughout much of the adenoma, with smaller areas of more widespread expression representing less than one third of the total area of the polyp in all cases (n = 3). The areas of more widespread FasL expression were well demarcated and coincided with areas of dysplasia within the polyps. The pattern of FasL expression within these areas was identical to that of adenomatous polyps, with more than 70% of epithelial cells expressing FasL.
Discussion
Our results show that FasL expression is upregulated at the adenoma stage of colonic carcinogenesis. Colonic adenomas expressed FasL irrespective of adenoma size, histological type, or the degree of dysplasia. Most cells in colonic adenomas coexpressed Fas and FasL without undergoing apoptosis. FasL expression by normal colonic epithelium and hyperplastic polyps was restricted to the luminal surface, where expression coincides with epithelial apoptosis. Serrated adenomas have a mixed pattern of FasL expression. Throughout most of their extent, FasL is expressed at the luminal surface, as in normal colonic epithelium and hyperplastic polyps. In areas of dysplasia within serrated adenomas, FasL expression is upregulated and expressed throughout these areas in the same pattern as is found in adenomatous polyps.
Apoptosis is a central feature of the regulation of colonic epithelial cell numbers.28 Currently, the mechanisms and control of apoptosis in colonic epithelial cell turnover are poorly understood. Although FasL expression was initially thought to be restricted to lymphoid cells, non-lymphoid FasL expressing cells include oesophageal epithelial cells,7 keratinocytes,8 Sertoli cells,9 ocular cells,10 cytotrophoblasts,11 and neurones.12 Fas is widely expressed in most cell types, including epithelial cells, and Fas mediated apoptosis might be involved in cell turnover in many tissues,14 including oesophageal epithelium,7 vaginal epithelium,15 the liver,16 and the testis.17 Our finding that terminally differentiated colonic epithelial cells at the luminal surface express FasL suggests a possible role for autocrine Fas mediated apoptosis in cell turnover.
Acquisition of resistance to apoptosis is thought to be crucial in the transformation of cells from normal to neoplastic in many tissues. A functional apoptotic pathway appears to be essential to maintain low numbers of mutated cells.29 Cells that have acquired resistance to apoptosis either before or as a result of mutational changes fail to be eliminated by apoptosis, and are thus able to continue the progression towards a neoplastic phenotype. Failure of apoptosis may be of particular importance in the development of colorectal cancer.30 There is in vitro23 and in vivo22 evidence that colon cancer cells are resistant to apoptosis mediated by Fas. Despite intracellular defects in Fas signalling in colon cancer cells,31 the Fas receptor is downregulated in many colonic carcinomas. Fas downregulation might be a consequence of p53 mutation at later stages of colon tumour development because Fas is a target gene for transcriptional activation by p53.32 Transfection of wild-type p53 into various tumour cell lines has been shown to restore/upregulate Fas expression.32
Our results demonstrate that Fas–FasL coexpression occurs in the absence of apoptosis in most epithelial cells in mildly dysplastic adenomas, a very early stage in the adenoma–carcinoma sequence. Fas–FasL coexpression without apoptosis was also seen in peritumoral epithelium. This suggests that colonic epithelial cells acquire resistance to Fas mediated apoptosis early in the adenoma–carcinoma sequence. Hyperplastic polyps retained the same pattern of FasL expression as the normal colonic epithelium, which is in keeping with their lack of well defined premalignant potential.
Finally, we previously demonstrated in vitro that FasL expression by colonic carcinoma cell lines may induce apoptosis of Fas sensitive immune effector cells.23 Evidence from human and animal studies supports this “Fas counterattack” mechanism of tumour immune escape in vivo.22, 33 Our finding that FasL is upregulated at the adenoma stage of colon carcinogenesis suggests that colonocytes acquire mechanisms of immune escape early during transformation.
Acknowledgments
We are indebted to J O'Callaghan and R Limmer for technical assistance. We are grateful to Dr G Lee, histopathologist, for advice, and to the staff and patients at the Mercy Hospital, Cork, for their cooperation. This research is supported by the Health Research Board of Ireland, the Wellcome Trust, the Cancer Research Appeal at the Mercy Hospital, Cork, Ireland, and the Irish Government Science and Technology Board (Forbairt).