Article Text

Abstract
Aims: Essential thrombocythaemia (ET) and primary myelofibrosis (PMF) share some clinical and pathological features, but show different biological behaviour and prognosis. The latest contributions to understanding the nature of these disorders have focused on bone marrow microenvironment remodelling and proliferative stress, recognising megakaryocytes (MKCs) as “key-cells”. The aim of this study was to investigate the apoptotic profile of ET and PMF MKCs in order to further characterise the biology of these disorders.
Methods: Bone marrow biopsy samples from 30 patients with ET, and 30 patients with PMF, were immunophenotypically studied for the expression of pro-apoptotic (Fas, Fas-L, Bax, Bad) and anti-apoptotic (Bcl-2, Bcl-XL, hTERT (human telomerase reverse transcriptase)) molecules and the “executioner” molecule caspase-3. The fraction of MKCs undergoing apoptosis was assessed by deoxynucleotidyl transferase-mediated dUTP nick-end labelling.
Results: Only the mitochondrial pathway seemed to be involved in MKC apoptosis. The anti-apoptotic molecule Bcl-XL was predominantly found in ET MKCs (50.5% of ET MKCs versus 35% of PMF MKCs; p = 0.036), while pro-apoptotic molecules Bax and Bad showed a prevalent expression in PMF MKCs (30.5% of ET MKCs versus 55% of PMF MKCs; 41% of ET MKCs versus 52% of PMF MKCs; p = 0.001 and p = 0.068, respectively). A significant fraction of PMF MKCs were committed to apoptosis according to caspase-3 expression and TUNEL, while only few ET cells were committed to apoptosis. hTERT was significantly more expressed in PMF (32% of ET MKCs versus 46% of PMF MKCs; p = 0.022), in agreement with the proliferative nature of this disease.
Conclusions: It was found that ET and PMF MKCs, which barely differ in terms of morphology and aggregation, are characterised by markedly different apoptotic profiles. The rather high apoptotic fraction of PMF was able to support the fibrotic nature of this process, while the anti-apoptotic profile of ET cells fits well with their “steady” maturative state.
Statistics from Altmetric.com
Philadelphia-negative chronic myeloproliferative disorders (Ph− CMPDs), namely essential thrombocythaemia (ET), polycythaemia rubra vera (PV) and primary myelofibrosis (PMF), derive from the transformation of a pluripotent bone marrow (BM) stem cell, resulting in the clonal proliferation of one or more myeloid lineages.1
In these disorders, thrombocythaemia can be the only appreciable clinical finding on presentation and can be complicated by thromboembolic and haemorrhagic episodes.
The differential diagnosis between Ph− CMPDs and reactive thrombocytosis is based on a set of clinical and pathological criteria. BM histopathology plays a relevant role in the diagnosis as defined by the World Health Organization classification.2 3
Although the identification of different forms of CMPDs in their full-blown stage is feasible, the correct classification of the early stages of CMPDs remains an issue of concern. In addition to well-stated histomorphological features mainly focusing on megakaryocyte (MKC) morphology, additional information may come from the assessment of the microvascular density, megakaryocyte proliferation index and morphometric analysis.4 5
The recent discovery of the JAK2 gain-of-function mutation has shed new light on the biological mechanisms underlying myeloproliferation in Ph− CMPDs.6 7 The JAK2 mutation has been found with different incidence rates in each of the CMPDs: more than 90% in PV, and up to 50% in ET and PMF.8 This represented the rationale for the introduction of JAK2 molecular defect evaluation in the diagnostic procedure of CMPDs in order to eliminate the need for further tests performed to exclude reactive myeloproliferative conditions.9 However, the existence of JAK2-negative cases and, among them, cases that do not show mutations in type-1 cytokine receptors and granulocytic colony stimulating factor receptors, suggests an involvement of pathways other than JAK2/STAT, at least in ET and PMF.8
The lack of clear-cut biological and histopathological differences between ET and early PMF is in contrast with the rather different clinical courses of the two diseases. While patients with ET do not commonly experience a relevant reduction of life expectancy, patients with PMF show a more progressive disease with less favourable outcome.
The latest contributions to the understanding of ET and PMF biology have focused on bone marrow microenvironment remodelling and yeloproliferative stress.10 In ET and PMF, a better comprehension of their biology can be achieved by taking into account the induction and regulation of apoptosis, as clonal expansion can be due to a decrease in programmed cell death in addition to a high rate of proliferation.11 Few in vitro studies investigating apoptosis in platelet and MKC differentiation have been reported so far.12 13
Apoptosis is mostly promoted by the means of an evolutionary conserved family of aspartic acid-specific cysteine proteases (caspases) that are normally present in the cell as latent precursors (pro-caspases).14 Caspases have been categorised into upstream initiators and downstream executioners, and they act through a proteolytic cascade leading to cell death. Initiator caspases include caspase-8 and caspase-9, which can be activated following different membrane or mitochondrial stimuli.14 The membrane (extrinsic) pathway involves cell surface receptors with an intracellular death domain such as CD95 (Fas), while the mitochondrial (intrinsic) pathway begins with the release of cytochrome c from mitochondria, and this is regulated by the Bcl-2 family of proteins.15 16
In the present study, we investigated the apoptotic profile of MKCs in ET and PMF by assessing the immunohistochemical expression of several pro-apoptotic and anti-apoptotic molecules involved in the intrinsic and extrinsic pathways of apoptosis.
PATIENTS AND METHODS
Patients
For this study, 60 bone marrow biopsies (BMBs) of patients with Ph− CMPD, performed on diagnosis and before starting any treatment, were selected from the archives of the Human Pathology Department of the University of Palermo. Ten cases of reactive thrombocytoses and five cases with normal histology or non-specific reactive changes were also included in the study as controls.
For the study, 30 cases of ET and 30 cases of PMF, which fully met the World Health Organization diagnostic criteria, were selected among those diagnosed between January 2002 and January 2005.
The following data were retrospectively gathered from the medical records of the patients, and are summarised in table 1: age, gender, white blood cell (WBC) count, red blood cell (RBC) count, platelet (PLT) count, haemoglobin (Hb), spleen size, presence of thrombotic risk factors, treatment, CMPD-related pathological events (occurring at presentation, or after diagnosis) and outcome.
Thrombotic risk factors were considered as follows.17 18 For ET: age >60 years, previous major thrombotic event and leukocytosis (WBC count above the median value); for PMF: thrombocytosis (PLT count above the median value), presence of any cardiovascular risk factor and Hb >11 g/dl.
Histopathological and immunophenotypical evaluation
Histological studies were based on Schaffer-fixed, paraffin-embedded BMBs using H&E, periodic acid–Schiff, Giemsa and Gomori staining.
Only representative bone marrow trephine biopsies (ie, non-tangential biopsies of more than 1.5 cm in length) collected at the time of diagnosis, before any treatment was started, were included in the analysis.
All BMBs were subjected to a histopathological re-evaluation, including the assessment of the amount of fibrosis according to the recently established semi-quantitative scoring system proposed in the European consensus on grading bone marrow fibrosis.19
Immunohistochemistry was performed according to the following procedure. Slides were heated in a microwave oven thrice for 5 min in TRIS/EDTA pH 9.0 buffer (Dako, Glostrup, Denmark), brought to room temperature, and washed in phosphate-buffered saline (PBS). Endogenous peroxidase activity was quenched for 10 min by H2O2 incubation. Following PBS washing, sections were incubated with protein block solution (Novocastra Laboratories, Newcastle upon Tyne, UK) for 10 min in a humidified chamber, and rinsed again in PBS. Primary antibodies were added to the sections at the correct concentrations and incubated for 1 h at room temperature in a humidified chamber. After washing three times in PBS , the immune complexes were detected by using the LSAB+ kit peroxidase (Dako) with 3-amino-9-ethylcarbazole. The following primary antibodies were used: mouse monoclonal anti-human Fas, 1:30 (Santa Cruz Biotechnology, Santa Cruz, California, USA); polyclonal anti-human Fas-ligand, 1:100 (Santa Cruz Biotechnology); monoclonal anti-human Bcl-2, 1:50 (Dako); mouse monoclonal anti-human Bcl-XL, 1:50 (Santa Cruz); mouse monoclonal anti-human Bax, 1:50 (Santa Cruz); mouse monoclonal anti-human Bad, 1:50 (Santa Cruz); mouse monoclonal anti-human hTERT (human telomerase reverse transcriptase), 1:20 (Novocastra Laboratories); polyclonal anti-human caspase-3, 1:120 (Promega, Madison, Wisconsin, USA).
The expression of each of the tested molecules in MKCs was assessed by counting the number of positive cells out of a total of 100 MKCs per section, and the result was expressed as a percentage. In the control cases (cases of reactive thrombocytosis, uninvolved marrows in staging procedures for lymphoma) the percentage of positive MKCs was calculated for 50 MKCs per section.
TUNEL assay
For the terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick end labelling (TUNEL) reaction, sections (4 μm thick) were permeabilised with 0.1% Triton X-100 in 0.1% sodium citrate for 2 min on ice. Apoptosis was determined by using the In Situ Cell Death Detection AP kit (Boehringer Mannheim, Indianapolis, Indiana, USA) according to manufacturer’s instructions. DNA strand breaks were detected by using 5-bromo-4-chloro-3-indolyl-phosphate (Dako) substrate. Positive controls were obtained by pretreating specimens with DNAse I (1 μg/ml) to introduce non-specific strand breaks, while the negative controls were performed using the TUNEL staining without TdT.
Apoptotic cells marked by TUNEL showed a dark-blue/black nuclear staining. The percentage of apoptotic cells detected by TUNEL was obtained as described for immunohistochemistry.
Statistical analysis
To evaluate differences in the immunophenotypical expression of pro-apoptotic and anti-apoptotic molecules among the ET, PMF and control groups, Mann–Whitney U test was performed; p values <0.05 were considered to be significant.
Initial clinical and laboratory features detailed in Patients and methods were analysed for their possible association with the apoptotic fraction of the cases studied. The χ2 test was performed, using Yates’ correction for continuity due to the small number of cases. Fisher’s exact test was used when one or more cells showed expected frequencies lower than five. A p value of 0.05 or less was considered to confirm associations with a probability of 95%. Computations were performed using the SPSS 13.0 software package (SPSS, Chicago, Illinois, USA).
RESULTS
Clinical presenting features and outcome
Clinical presenting features of the patients studied are summarised in table 1. Overall, 24 patients with ET (80%) and 23 with PMF (76%) were asymptomatic at the time of diagnosis. Twelve patients with ET (40%) and 21 patients with PMF (70%) were classified at high-risk of thrombosis at the time of diagnosis. Adverse pathological events related to the underlying myeloproliferative disease occurred in the form of thrombotic events in eight patients with ET (26%) and 10 patients with PMF (33%), respectively. All the patients with ET, and all but one of the patients with PMF who underwent thrombotic complications were high-risk patients. With a median follow-up times of 39 and 36 months for ET and PMF groups, respectively, only one death was recorded in the PMF group and it was due to causes unrelated to the CMPD.
Histopathological and immunophenotypical evaluation
All the original diagnoses were confirmed on histopathological re-evaluation and the PMF cases were classified according to the degree of fibrosis: four cases were scored PMF-0, 21 were scored PMF-1, and five were scored PMF-2.
The status of activation of the apoptotic pathway in ET and PMF was studied by investigating the immunophenotypical expression of several pro-apoptotic and anti-apoptotic molecules involved in the membrane and mitochondrial pathways of apoptosis. Their expression was also assessed on MKCs from cases of reactive thrombocytosis and from “normal” bone marrows.
A schematic representation of the two apoptotic pathways, including the molecules studied, is given in fig 1, while table 2 summarises the results that were obtained by the assessment of the number of positive MKCs. The statistical analysis was performed only for those markers found to be expressed in MKCs.
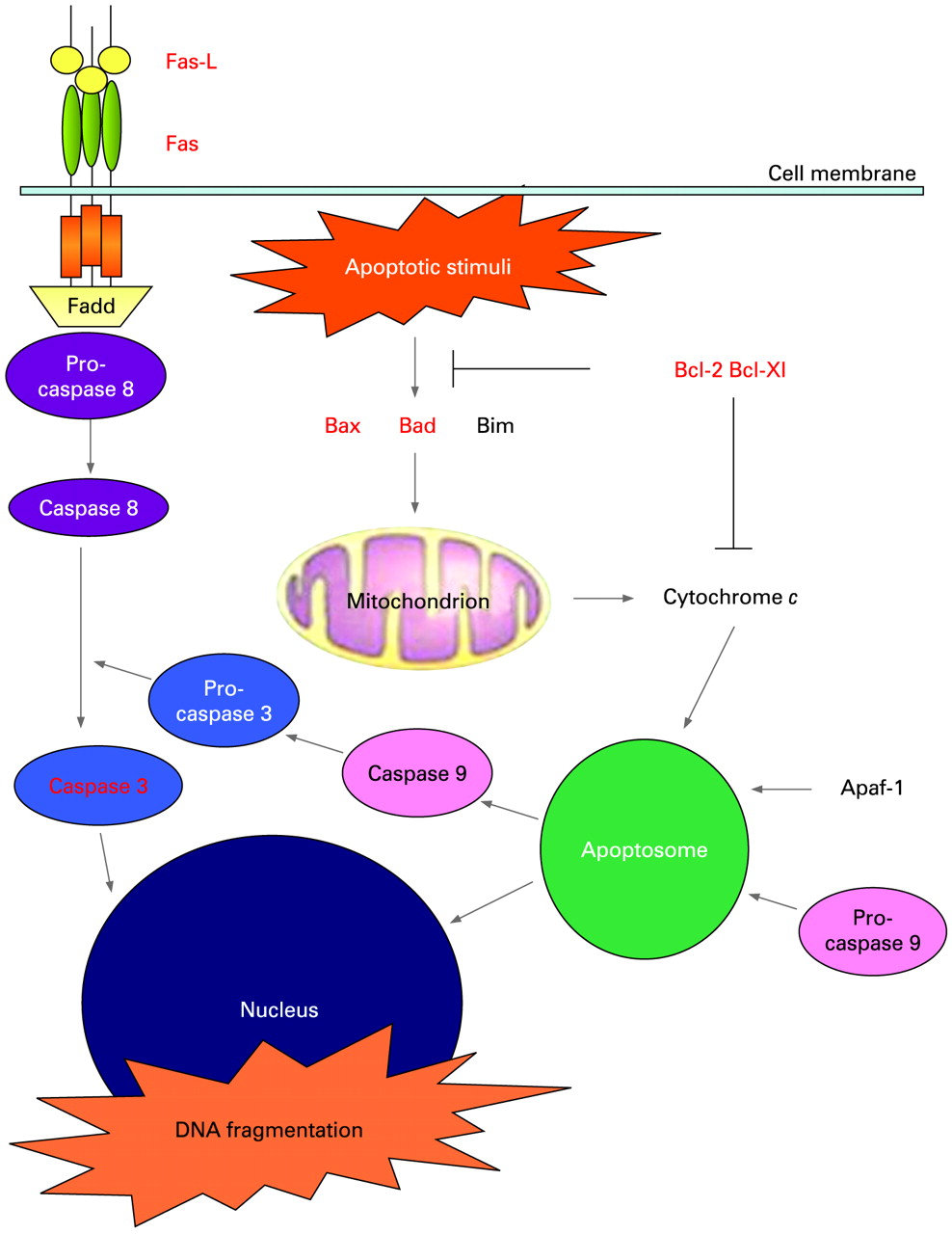
Schematic representation of the membrane (extrinsic) apoptotic pathway involving cell surface receptors with intracellular death domain (Fas) and of the mithocondrial (intrinsic) pathway starting with the release of cytochrome c from mithocondria regulated by Bcl-2 family proteins. The molecules investigated are shown in red.
CD95 (Fas), a receptor playing a role in the membrane pathway, was not expressed on the surface of MKCs from CMPDs or controls, while its ligand Fas-L was detected on the surface of few MKCs from control samples. The only sign of Fas-L expression in MKCs from CMPDs was observed around the surface of leukocytes undergoing emperipolesis (fig 2F).

(A, B) A larger number of megakaryocytes (MKCs) stain positive for Bcl-XL in essential thrombocythaemia (ET) (A) than in primary myelofibrosis (PMF) (B). Bcl-XL, StreptABC, original magnification ×250. (C, D) Only a small fraction of MKCs express Bax in ET (C) compared with PMF (D). Bax, StreptABC, original magnification ×250. (E) Bad shows a cytoplasmic dot-like reactivity in PMF MKCs. Bad, StreptABC, original magnification ×250. (F) Fas-L is detected inside PMF MKCs at the site of leukocyte emperipolesis (arrow). Fas-L, StreptABC, original magnification x400.
Among the anti-apoptotic molecules involved in the mitochondrial pathway, Bcl-2 was not expressed in MKCs from CMPD and control samples. On the other hand, Bcl-XL was found in less than 5% (SD 2.88) of MKCs from control samples.
In CMPDs, a higher percentage of MKCs positive for BCL-Xl was observed in ET (mean 50.5%, SD 24.19) than in PMF (mean 35%, SD 28.06) cases (p = 0.036). MKCs expressing BCL-Xl showed a diffuse cytoplasmic pattern of positivity (fig 2A, B).
As regards Bax and Bad, which are molecules with pro-apoptotic function acting in the same mitochondrial Bcl-2-related pathway, the following percentages were observed in control samples: Bax, mean 17.5%, SD 12.05; Bad, mean 30%, SD 15.28. In ET a modest fraction of MKCs (mean 30.5%, SD 19.87) expressed Bax in the form of a diffuse cytoplasmic reactivity while in PMF cases it was expressed by the majority of typical MKCs (mean 55%, SD 18.89) (p = 0.001) even if some negative cells were found (fig 2C, D). Similarly Bad showed a prevalent expression in PMF MKCs (mean 52%, SD 25.20) compared with ET MKCs (mean 41%, SD 24.18, but the difference was not significant, p = 0.068). In MKCs Bad showed a dot-like mitochondrial pattern of staining (fig 2E).
The effective activation of the apoptotic process in its fully developed phase was assessed by means of the detection of the cleaved form of caspase-3. In PMF, a significant fraction (29.5%, SD 23.51) of MKCs committed to apoptosis was observed. The same dot-like pattern of expression was found also in ET cases, where only few cells were positive (mean 11%, SD 15.41) (p = 0.006). A percentage of caspase-3-expressing MKCs similar to that observed in ET cases was found in controls (mean 13%, SD 7.32) (fig 3A, B).

{kind=link}
{kind=link}
{kind=link}
(A, B) Active caspase-3 is expressed in the form of a clear dot-like cytoplasmic pattern in most of the primary myelofibrosis (PMF) (B) and a few essential thrombocythaemia (ET) (A) megakaryocytes (MKCs). Active caspase-3, StreptABC, original magnification ×250. (C, D) Deoxynucleotidyl transferase (TdT)-mediated dUTP nick end labelling (TUNEL) assay shows a higher number of apoptotic MKCs undergoing DNA fragmentation (black nuclei) in PMF (D) than in ET (C). Apoptotic figures are also present in the context of intrasinusoidal haemopoiesis (inset). TUNEL staining, original magnification ×400.(E, F) hTERT (human telomerase reverse transcriptase) is expressed in MKCs with a nucleus-restricted pattern. The percentage of positive cells is greater in PMF (F) than in ET (E). hTERT, StreptABC, original magnification ×400 (E), ×250 (F).
The actual apoptotic fraction of MKCs was visualised also highlighting DNA fragmentation by means of the TUNEL assay. The reaction confirmed the data for caspase-3 although the percentages of positive MKC nuclei were lower in PMF (mean 22.5%, range 5–40%) and ET (mean 9%, range 0–20%) (fig 3C, D). Apoptotic figures were also observed in intrasinusoidal haematopoiesis, which is a typical feature of PMF (fig 3D, inset).
The expression of hTERT was investigated in order to assess telomerase activity. In MKCs from control samples, as well as in MKCs from cases of reactive thrombocytosis, no expression of hTERT was detected.
In ET and PMF MKCs, the expression of hTERT was restricted to the nucleus; the percentage of positive cells was greater in PMF (mean 46%, SD 17.77) than in ET (mean 32%, SD 15.52) (p = 0.022) (fig 3E, F).
Clinical and pathological correlations
Following immunophenotypical analysis, we wanted to test for associations between the MKC apoptotic fraction of ET and PMF cases and the clinical laboratory and follow-up data.
On statistical analysis, none of the presenting clinical and laboratory features was found to be associated with the apoptotic fraction except for the presence of thrombotic events. Actually, cases with a high Bcl-XL expression (⩾80%) and a low apoptotic fraction (caspase-3 ⩽5%) proved to be significantly associated with the occurrence of pathological thrombotic events (p<0.001) in ET (six out of eight cases, 75%) and PMF (seven out of ten cases, 70%) groups. All these cases with a low apoptotic profile, and in which a thrombotic event was recorded, showed at least one risk factor for thrombosis at the time of diagnosis. Although a positive association between the presence of one or more risk factors and the occurrence of pathological events was also observed (p = 0.02), there was no significant association between risk factors and the apoptotic fraction.
DISCUSSION
The need for an early and careful discrimination between ET and pre-fibrotic PMF, which are two disorders with a similar clinical and morphological picture but different outcome, has stimulated the search for criteria aimed to delineate more precise diagnostic categories. The bone marrow histopathological approach for CMPDs focuses on the evaluation of an array of differential features that include bone marrow cellularity, MKC morphology and aggregation, myeloid metaplasia, fibre content and the evaluation of non-megakaryopoietic parenchyma, and it identifies MKCs as “key cells”.20 In ET, as well as in pre-fibrotic and early-fibrotic PMF, the clonal proliferation seems to involve primarily MKCs, which barely show differences in terms of morphology, morphometry and aggregation.4 5 Immunophenotypical findings such as microvessel density, and presence and number of CD34 cells and proliferating MIB-1, cells have all been taken into consideration thus indicating the leading role of immunohistochemistry in diagnostic and prognostic evaluation.4 21 22 Together, the aforementioned discriminative features allow a correct diagnosis in most cases, but they are not sufficient to explain the different biological background of these disorders.
The most recent and significant contribution to this issue has been achieved by the identification of JAK2 gain-of-function mutations (V617F, exon 12), which are likely to be responsible for proliferative stress maintenance, although the pathogenetic role of the JAK–STAT pathway in the development of CMPDs is not yet fully understood.8
To assess whether the activation status of the apoptotic pathway in ET and PMF is different, we tested the immunophenotypical expression of several pro-apoptotic and anti-apoptotic molecules involved in the membrane and mitochondrial pathways of apoptosis.
The extrinsic apoptotic pathway did not appear to be involved in MKC apoptosis, as neither CD95 nor its ligand Fas-L were found significantly expressed on the surface of MKCs from CMPD samples. The exclusive expression of Fas-L on leukocytes undergoing emperipolesis is in agreement with the fact that this pathway is driven by a microenvironmental death signal followed by the ligation of membrane death receptors.23
Actually, the expression of Fas-L and other tumour necrosis family member ligands on activated platelets has been described, and its reactivity on phagocytosed cells could follow MKC activation.12
Relevant differences were observed in the expression of molecules involved in the intrinsic pathway. This pathway is initiated when repair mechanisms cannot cope with internal cell injuries, and it is characterised by an imbalance between pro-apoptotic and protective molecules regulating mitochondrial permeability.11
Among the anti-apoptotic molecules acting in this pathway, Bcl-2 expression was not found either in MKCs from CMPDs or in control samples. Conversely, MKCs displayed Bcl-XL positivity in control and cases of CMPD, and in ET cases a higher percentage of positive MKCs was detected as compared with cases of PMF. The evidence of a more prominent fraction of Bcl-XL-expressing MKCs in ET than in PMF could seem to be in contrast with recent in vitro observations reporting an early Bcl-XL downregulation during ET MKC differentiation13 and a significant expression of Bcl-XL in PMF MKCs.24 Actually, those two studies relied on thrombopoietin-cultured MKCs differentiated from CD34 peripheral blood cells. Data coming from such in vitro models should therefore be interpreted with caution, as in vitro cultured cells lack stimuli coming from cytokines, growth factors, cell–cell and cell–BM–stroma interactions which are well known to influence their proliferation and survival in vivo. However, the broad range of variation in Bcl-XL expression we observed in our cases of ET and PMF does not indicate its usefulness as a diagnostic feature for differentiating ET from PMF.
Pro-apoptotic molecules, such as Bax and Bad, acting in the same mitochondrial Bcl-2-related pathway, behaved differently in that both showed higher levels of expression in PMF MKCs than in ET MKCs. Bax and Bad are multidomain pro-apoptotic Bcl-2 family proteins, which upon activation exert their pro-apoptotic activity by facilitating the release of cytochrome c and the sequential activation of the caspases.25 It has been recently demonstrated that an increase in Bax, even in the absence of Bad, is able to trigger the apoptotic process.26 In a study analysing gene expression profiles of BM CD34-derived MKCs from patients with ET and from healthy subjects, Tenedini et al.27 found that Bax, along with other pro-apoptotic genes such as BNIP3 and BNIP3L were downregulated in ET MKCs whereas anti-apoptotic genes (like IGF1-R and CFLAR) were upregulated. Their conclusion, after in vitro characterisation of apoptosis of normal and ET MKCs, was that ET MKCs are more resistant to apoptosis than their normal counterparts; this is in agreementwith our data, even though the extension of such observations to in vivo MKCs deserves caution.
The actual fraction of cells committed to apoptosis was studied analysing the expression of the effector caspase-3 and confirmed by means of TUNEL assay. The different expression of caspase-3, in the two disorders is in keeping with the higher proportion of MKCs reaching the execution phase of the apoptotic process in PMF, consistently with the significant expression of pro-apoptotic molecules. The smaller fraction of positive MKCs observed by TUNEL can be ascribed to the fact that this assay highlights DNA fragmentation occurring late in apoptosis.
In order to evaluate the response of CMPD MKCs to cellular ageing, we also investigated telomerase activity by means of hTERT expression. hTERT is the catalytic subunit of telomerase, which is a reverse transcriptase that is not usually found in resting somatic cells. It allows the overcoming of the mitotic checkpoint through telomere elongation but does not prevent the cell from dying owing to pro-apoptotic stimuli external to the replicative cycle.28
In our study hTERT was expressed at higher levels in PMF MKCs than in ET MKCs and, although the significant fraction of hTERT positive MKCs observed in IMF seems to be in contrast with the pro-apoptotic profile delineated so far, it actually it fits well with the overt proliferation rate of MKCs in PMF.
In a study based on the quantitative evaluation of this molecule in myelodysplastic syndromes and CMPDs, Bock et al29 found that it was much more expressed in the former and that no significant differences were observed among the different CMPDs. The different expression of hTERT we observed in the two CMPDs studied could be due to the fact that our analysis was limited to the megakaryocytic population, and did not take into account the non-megakaryocytic parenchyma.
An interesting finding, although very preliminary due to the small number of cases and the non-clinical approach of the study, was that patients undergoing thrombotic events related to the underlying CMPD showed the lowest fraction of apoptotic MKCs in ET and PMF groups. If confirmed, this evidence would suggest that cases with a prominent anti-apoptotic phenotype can be characterised by a more “active” disease being more prone to thrombotic complications.
It has also been suggested that the occurrence of thrombotic and haemorrhagic events can be predicted from the biological characteristics of the disease, other than the presence of markers of hypercoagulability and general cardiovascular risk factors.30 Among the former, besides an impaired expression of c-mpl in bone marrow MKC,31 a recent study provided evidence of how the rate of thrombotic complications in JAK2-positive patients with ET was significantly higher than that in wild-type patients with ET and not statistically different from that in patients with PV.32 As in our series, JAK2 mutational status was assessed in a low percentage of cases (20%), and its association with the occurrence of adverse events cannot be evaluated. Nevertheless, the relationship between JAK2 and the apoptotic profile would be worth studying.
In conclusion, we found a higher percentage of MKCs committed to apoptosis in PMF compared with ET, and this was supported by a different expression of pro-apoptotic and anti-apoptotic molecules in the two disorders; however, the high degree of variation we observed in the expression of such molecules does not allow their use for diagnostic purposes. According to our evidence, ET MKCs displayed a non-proliferative, non-apoptotic “steady” profile that fits perfectly with the term “over-aged”, while the profile of PMF MKCs was that of actively proliferating cells with a possibly shorter life span. The relatively high fraction of apoptotic cells in PMF could be related to the fibrotic nature of this disease, as apoptotic cells and their debris can constitute a fibrogenic spur.
Take-home messages
Essential thrombocythaemia (ET) and primary myelofibrosis (PMF) are tightly linked by the prominent involvement of megakaryocytes (MKCs) in myeloproliferation. However, the MKC populations from these two disorders differ significantly on a biological basis.
Here we show that PMF MKCs display a phenotype of cells prone to proliferation and apoptosis different from that of ET MKCs, which show prevalent expression of anti-apoptotic molecules.
Together these results contribute to clarify the issue of the biological diversity of ET and PMF, further supporting the myeloaccumulative nature of the former and the progressive behaviour of the latter.
REFERENCES
Footnotes
Competing interests: None.