Article Text

Abstract
Aims Grafts have been shown to be sites where the alloimmune response develops in a direct interaction between the targeted tissue and the immune effectors. An important issue in renal rejection is B cell infiltrate that may contribute to the development or persistence of rejection. Analysis of gene-expression patterns also provides a window on the biology and pathogenesis of renal allograft rejection.
Methods To better understand the role exerted by B cells in a renal acute rejection, the authors analysed the IgVH gene repertoire in six cases of transplanted kidneys with acute T cell-mediated rejection (TCMR), three of which were associated with antibody-mediated rejection (ABMR).
Results The authors found mutated and unmutated sequences, without any evidence of clonal relationships, in all patients with TCMR alone and in two of the three cases with both acute TCMR and ABMR. The remaining patient showed glomerular inflammation and thrombosis, with diffuse C4d glomerular and peritubular capillary deposition, and hypermutated V region genes.
Conclusions These results suggest that there is more than one pathway to the onset and perpetuation of CD20 (+) B cells infiltration in acute rejection; furthermore, the CD20 (+) B cells' clonal expansion may be responsible for a more severe pattern of ABMR, through immune-mediated tissue damage.
- T-cell-mediated rejection
- antibody-mediated rejection
- IgVH gene analysis
- immunoglobulin
- kidney
- rejection
- transplantation
Statistics from Altmetric.com
- T-cell-mediated rejection
- antibody-mediated rejection
- IgVH gene analysis
- immunoglobulin
- kidney
- rejection
- transplantation
Introduction
The involvement of antibodies in rejection of organ transplants is increasingly recognised as a relevant pathomechanism. In acute cellular liver allograft rejection, hepatic B lymphocytes and plasma cells as well as B-cell-activating cytokines have recently been described, indicating that, in addition to T cells, antibody-mediated mechanisms might be involved. It is well known that the success of kidney transplantation is dependent on the genetic and immunological compatibility of the grafted organ with the recipient.
Biopsy samples from patients with acute kidney allograft rejection that are indistinguishable on conventional histological analysis reveal extensive differences in gene expression, which are associated with differences in immunological and cellular features and clinical course. It is therefore amazing that, in renal rejection, there are few data on the stage of differentiation of B cell infiltrates and their role in the development or persistence of rejection.1–10
The analysis of the immunoglobulin variable heavy chain region (IgVH) genes had a great impact on the understanding of B-cell response in various inflammatory and autoimmune diseases but had never been applied to kidney allograft rejection.11–17 It has been applied to acute humoural and cellular liver allograft rejection, suggesting that locally accumulated B cells and their antibodies might be involved in IgG-mediated complement activation.18
Thus, our aim was to analyse the IgVH gene repertoire in transplanted kidneys in six cases of acute T cell-mediated rejection (TCMR), three of which were associated with antibody-mediated rejection (ABMR) in order to better understand the role played by CD20(+) B cells in renal acute rejection.
Materials and methods
Patients and tissue
Percutaneous renal needle biopsy guided by ultrasound (G18) was obtained from six recipients of deceased donor first kidney transplants performed at the Siena University Hospital between 2004 and 2006. The clinical data are resumed in table 1. The clinical course was characterised by suspected early acute rejection. The time of biopsy ranged from 15 days to 20 months after transplant.
Clinical data
Patients 1, 2 and 3 showed the presence of anti-HLA antibodies, revealed by the Luminex technique performed at the time of biopsies.
Renal biopsies met the Banff criteria for adequacy19; we used the Banff 2007 classification for allograft pathology and score.20 All biopsies were interpreted by two pathologists with expertise in renal allograft pathology.
Immunohistochemical stains were performed on 3 μm thick sections of each block employing the streptavidin–biotin method. After being dewaxed and rehydrated, sections were incubated with 3% H2O2 in Tris buffered saline (TBS) to inhibit endogenous peroxidase, then processed with different methods for each antibody. In order to show CD4(+) the sections were unmasked with Wcap buffer (pH 6.0 for 40 min at 98°C; Bio-Optica, Milan, Italy) and incubated with antihuman CD4 monoclonal antibody (clone 4B12; 1:50; Menarini, Florence, Italy). To improve the detection of CD8, CD20, CD27, CD68 and Ki67, we performed a pretreatment in citrate buffer (0.01 M, pH 6.0) with a microwave oven at 750 W for 5 min, for three cycles; sections were incubated with antihuman-CD8 monoclonal antibody (clone CD8-144B; 1:50; Dako, Milan, Italy), CD20 (clone Ab-1 (l26); 1:150; Neo Markers, Freemont, California), CD27 (clone 137B4; 1:50; Novocastra, Newcastle, UK), CD68 (clone KP1, 1:50; Dako) and Ki67 (clone SP6, 1:200; NeoMarkers, Freemont, California, USA). For CD38 (clone38C03; 1:50; Thermo Scientific, Fremont, California), slides were preincubated with EDTA buffer (0.05 M, pH 8.0) for 30 min at 98°C. For CD21 (clone 1F8, 1:10, Dako), the immunoperoxidase technique was performed after antigen retrieval with protease 0.4% in Tris buffered saline (TBS) buffer for 10 min at 37°C. The biotinylated secondary link universal antibodies were associated with Streptavidin-horseradish peroxidase (HRP, Dako Cytomation, Milan, Italy) and revealed with DAB (Dako Cytomation LSAB+System-HRP). For C4d, pretreatment with citrate buffer (0.01 M, pH 6.0) in a microwave oven at 750 W for 5 min (three cycles) was followed by incubation with antihuman C4d polyclonal antibody (1:100; DBA, Milan, Italy). The Primary Antibodies Enhancer was associated with AP Polymer (UltraVision LP Detection System AP Polymer, LAB Vision) and revealed with fuchsin (Dako Cytomation).
Known positive controls (tonsil tissue) as well as negative controls (without primary antibody) were also stained in each run.21 The monoclonal antibodies and source used are shown in table 2.
Panel of antibodies used for diagnosis
Histology
CD20(+) B cell infiltrate was distinguished in nodular and scattered distribution; nodular when clear dense clusters of CD20(+) B cells were present and scattered when at least a moderate (++) number of CD20(+) B cells were present in the interstitial compartment.
In either the cortex or the medulla, the staining for C4d in peritubular capillaries (PTCs) and scoring of peritubular capillaritis (PTCs) were evaluated as recommended in the Banff 2007 classification.20
Monocyte macrophages (CD68 (+), evaluated in glomeruli, PTCs and interstitium), T cells (CD3 (+), CD4 (+) and CD8 (+)) and plasma cells CD38 (+) were graded using a semiquantitative score (0=scanty to 3=conspicuous).
Laser capture microdissection
Clusters of CD20 (+) B cells were selected for microdissection from diffuse and nodular areas of B cell infiltrate. LCM was performed on 3 μm thick immunostained tissue taken from the previously described formalin-fixed paraffin embedded tissue.20 Microdissected cells from the two different areas were isolated and transferred to a PCR test tube.
Clusters and isolated T cells were identified by their immunophenotype and served as negative controls. The cells were dropped in 10 μl of PCR buffer (50 mM KCl; 10 mM Tris-HCl; pH 8.4; 0.01% gelatin) containing 200 μg/ml of proteinase K (Qiagen, Hilden, Germany). The isolated cells were covered with 50 μl of mineral oil and digested at 37°C for 16 h.21
PCR, cloning and sequence analysis
To avoid cross-contamination, all procedures prior to PCR amplification were performed in a room dedicated exclusively to this purpose and separated from the rooms where the subsequent steps were carried out. DNA quality control PCR, assessed by control gene primer sets according to the BIOMED 2 protocol,22 showed that all six gave an amplification of 300 bp or more and thus sufficient intact DNA for successful amplification of the IgVH rearrangement.
To identify the rearranged segments of the variable heavy chain genes in cell clusters (CD20 (+) cells) an FR2BM PCR method (according to the BIOMED 2 protocol) and a nested PCR method were performed. In the FR2BM PCR, the set of primers consisted of seven oligonucleotides capable of annealing to corresponding VHFR2 segments (VH1FR2–VH7FR2). These VH primer sets were used in conjunction with a single JH consensus primer, designed to anneal to the most homologous 3′-end of the six JH segments. For the nested PCR method, in the first round of amplification a mix of six VH family specific framework 1 (FR1) primers and a consensus JH primer (LJH) were used. For reamplification, an aliquot of the first PCR from each case was amplified with a family-specific VH primer set for framework 2 (FR2) and a nested JH primer (VLJH).
PCR conditions and primers have been described in detail elsewhere.23–25
The purified amplicons were cloned to the pCRII-TOPO vector (TOPO TA Cloning Kit; Invitrogen, Carlsbad, California) and were used to transform TOP 10 cells (Invitrogen) according to the manufacturer's instructions. A mean of at least 30 clones per case was sequenced in both directions with an ABI PRISM 310 Genetic Analyser (Applied Biosystems, Foster City, California). Sequences were compared against the GeneBank database (http://imgt.cines.fr) and the IgBlast database (http://www.ncbi.nlm.nih.gov/igblast/).25 VH genes were considered mutated if they differed by ≥1% from the corresponding germline sequence. In addition, sequences that showed the same V, D, J, identical CDR3 and the same pattern and number of somatic mutations were considered as one. Finally, family trees were constructed by analysing shared and unique mutations occurring in sequences that were clonally related.26 27
Two methods were applied to assess antigen selection in productive and mutated VH gene immunoglobulin rearrangements as described previously.21 28 In other types of calculations, as described previously,23 the calculation of a reliable p value (p≤0.05) failed in the cases with a very low number of somatic mutations.
Results
The patients' characteristics are summarised in table 1.
The clinical course of each patient was characterised by delayed function of the followed by early suspected acute rejection. Patients 1 and 4 showed graft failure with graft explantation.
The histopathological data for each case are shown in table 3.
Histopathological data
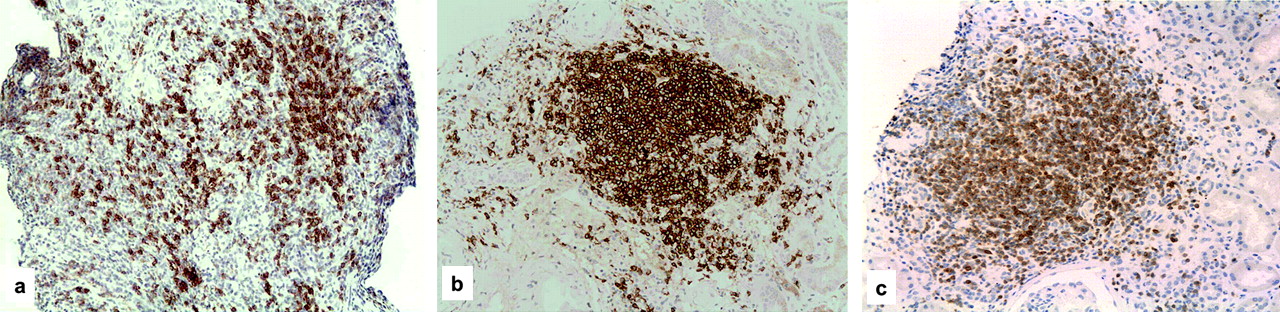
In all cases there was a scattered and nodular CD20 (+) B cell infiltrate (figure 1A,B). Some proliferating lymphocytes and a weak meshwork of CD21 dendritic cells were also detected in the nodules, but no clear germinal centre structures were identified; B-cells were intermingled with a minor component of CD4(+) T cells, and macrophages and plasma cells were present in the interstitium, but represented less than 10% in three cases. CD27 (+) B cells were scattered in the lymphoid infiltrates. C4d deposition was diffusely present in the PTCs in case 1 and focally present in cases 2 and 3. It is worth noting that glomerular capillary endothelial deposition of C4d (figure 1C) and glomerular inflammation with thrombosis and fibrinoid necrosis were only present in case 1 (figure 2).

CD20(+) B cell pattern of distribution. (A) Scattered CD20(+) B cell infiltrate; (B) nodular CD20(+) B cell infiltrate; (C) CD4+ T cells in nodules (immunohistochemical stain, (A, B): × 100; (C) ×200).

Histological picture of antibody-mediated rejection in case 1. (A) capillary thrombosis and cellular infiltration (H&E, ×200); (B) capillary fibrinoid necrosis (Masson Trichromic ×200); (C) diffuse capillary deposition of C4d (immunohistochemical stain ×100).
A total of 183 VH sequences were amplified from the six cases of kidney allograft analysed. Eighteen of the 139 rearranged VH genes (10%) were non-productive, being out of frame in five cases and containing a premature termination codon in CDR2 and CDR3 in three cases.
Of the nine potentially productive VH rearrangements amplified from CD20 (+) B cells, six were unique, and three were shared by different sets of cells (see below).
Sequence analysis of the productive rearrangements of CD20 (+) B cells showed that they represented a mixed population of mutated and unmutated CD20 (+) B cells, with a great variability between cases. The average mutation frequency range in mutated cells was 4.11–21.55% and did not differ significantly among the various clinical and pathological categories of renal transplant (table 4).
Molecular analysis of IgVH genes
Overall, the use of IgVH genes in our cases did not appear to show any bias towards the preferential use of a specific gene, since VH3 (the largest and most frequently used family) was found to be rearranged most often (108/139; 78%) followed, in order, by VH4 (20/139; 14.5%), VH5 (8/139; 6%), VH2 and VH1. The use of JH segments also appeared to reflect the normal representation of peripheral antigen-experienced CD20 (+) B cells.
The estimation of antigen selection in CD20 (+) B cells showed that the mutation pattern in most of their IgVH genes was negative for antigen-driven selection of a functional CD20 (+) B cell receptor. In fact, of 84 mutated IgVH genes, only 41 had an R/S ratio in the framework region (FWR) 3 <1.5 (11 out of 34 had an R/S ratio in the CDR2 >2.9) (table 4).
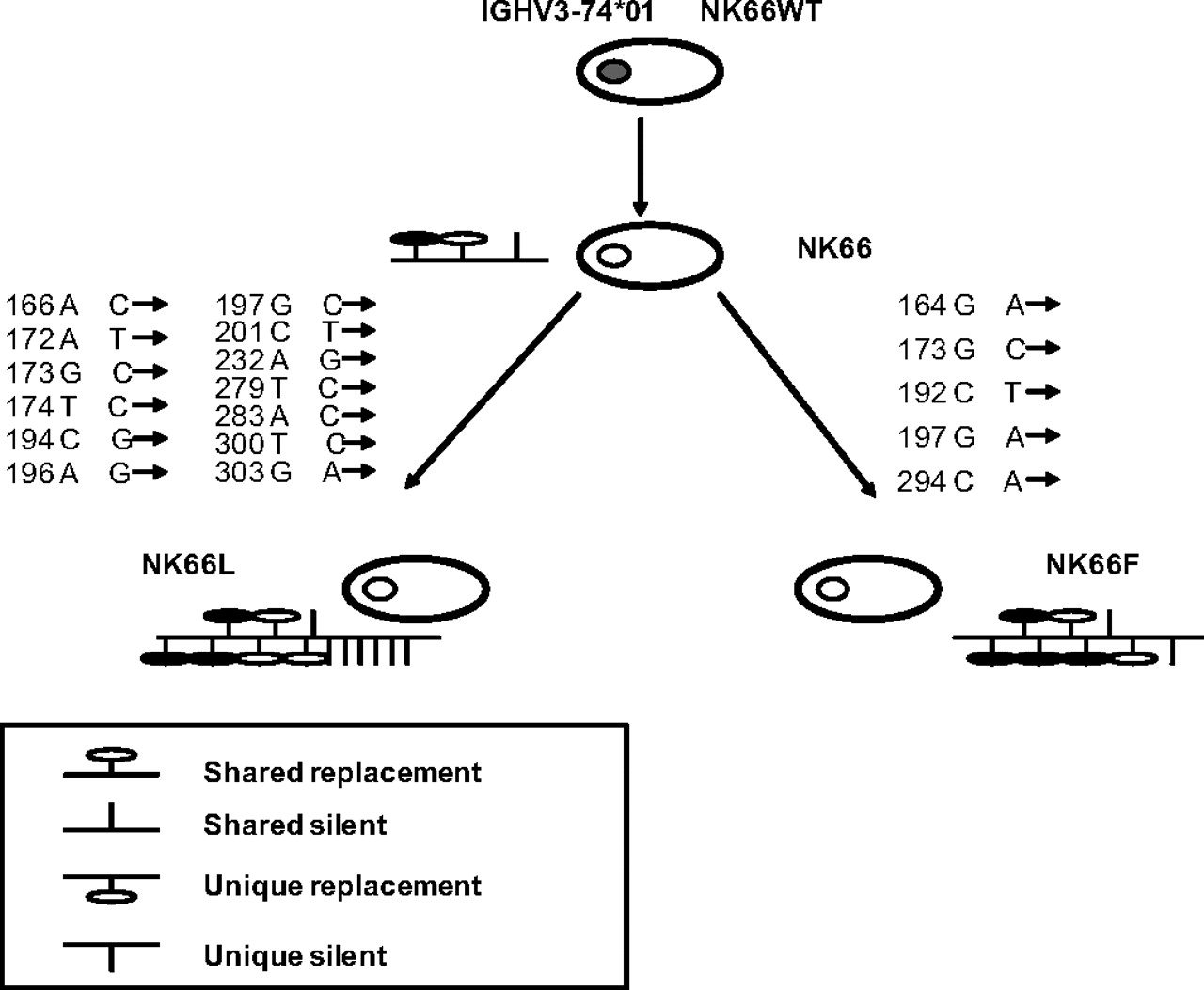
Regarding the three non-unique VH gene rearrangements amplified from case 1, all were productive and mutated and belonged to one group of three clonally related cells (clones L and F1). All three sequences showed a VH3-74*01/DH4-23*01/JH4*01 usage but a different pattern of somatic mutations. These clones had virtually identical CDR3s and carried shared as well as unique somatic mutations, suggesting clonal expansion and intraclonal variation (figure 3). In the same case, sequences with an identical CDR3 rearrangement but a different VH usage were isolated; in fact, the sequence derived from clone A had a VH3-15*05/DH5-24*01/JH4*01 rearrangement, whereas the sequence derived from the clone B1 had a VH4-59*08/DH5-24*01/JH4*01 rearrangement.

Clonal relationship between CD20(+) B cells in case one. The group of clonally related cells was composed of three cells obtained from three different single CD20(+) B cells. The putative progenitors are depicted with grey nuclei, and the IgV gene rearrangement of the clonally related cells deriving from the progenitor is indicated above. Shared point mutations and acquired unique mutations are indicated above and below the line, respectively. Vertical bars depict S mutations, and lollipops depict R mutations.
This finding indicates that the CD20 (+) B cells analysed in our cases had revised the receptor specificity (ie, receptor revision). The clones listed in figure 4 illustrate the VH replacement occurring at the 3′ end of the FR3 embedded heptamer (TACTGTC). The 3′ FR3 embedded heptamer is displayed with a black background in the figure, and HCDR3s are boxed.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
DNA clones exhibiting variable heavy chain (VH) replacement at the 3′ FR3 embedded heptamer. Two VH (VH 5–15, VH 4–59) clones from NK66 (A and B1) are aligned centrally with their most similar germline gene counterparts. The 3′ FR3 embedded heptamer is displayed with a black background, and the HCDR3s are boxed. Nucleotide differences between the two clones are indicated by asterisks, and identities are indicated by dots.
Note that the HCDR3s of the two clones are identical, whereas the VH segments upstream of the heptamer differ greatly between the two clones and resemble different germline genes.
Discussion
Characterisation of the B cell population in acute rejection is fundamental for a better comprehension of the role exerted by these cells in graft tissue, as well as for a correct therapeutic approach.8 9 The immunohistochemical evaluation of CD20(+) B cells infiltrates in allograft biopsies has been suggested for potential treatment with anti-CD20 mAb (rituximab),29–31 as the presence of CD20(+) B cell infiltrates and CD38(+) plasmablasts in the kidney biopsies of patients undergoing acute renal allograft rejection has been reported to be associated with glucocorticoid resistance and a high risk of graft failure.1 8 9 32–40
In our cases, we found lymphoid infiltration without any correlation with the clinical outcome. This is in line with recent reports that exclude any correlation of B cell infiltrates with the outcomes.10–12 32
Analysis of the IgVH gene repertoire has had a great impact on the understanding of CD20(+) B cell responses in lymphatic organs and has subsequently been applied to CD20(+) B cells in various inflammatory and autoimmune diseases and liver transplants but had never been applied to kidney allograft rejection.15 18 41
Our analysis of the VH-gene repertoire in two patients with both acute TCMR and ABMR and three patients with only TCMR showed the presence of mutated and unmutated sequences without any evidence of clonal relationships. This finding indicates that CD20 (+) B cells become activated in peripheral lymphoid organs and subsequently migrate into renal tissue.42 43 Therefore, in these cases the small aggregates of B lymphocytes are the result of local accumulation rather than the consequence of local CD20 (+) B cells activation and differentiation. On the other hand, the finding of two identical rearrangements, which differed from each other by up to 13 somatic mutations in case 1, speaks for antigen activation of B-cells and a clonal expansion within the renal tissue. The sequences allowed the construction of genealogical trees, which showed the stepwise accumulation of single somatic mutations during clonal expansion in the renal tissue (figure 3). The finding of mutated and clonally related VH genes in the lymphatic aggregates in case 1 seems to be in contrast with the absence of GC or GC-like structures in the same case, based on the assumption that B cells undergo somatic hypermutation in the GC stage of differentiation.
However, recent evidence in animal models has revealed that normal B cells in extrafollicular areas of secondary lymphoid tissue are capable of responding to antigens and undergoing clonal expansion. Interestingly, in the same case, we were also able to demonstrate evidence of receptor revision, suggesting a pressure for the antigen in this context. The possible occurrence of secondary rearrangement of Ig genes in peripheral lymphoid tissues remains controversial, although several studies have reported the existence of this phenomenon.42 43 In particular, receptor revision of VH genes has recently been proposed in both malignant lymphomas and normal human B lymphocytes.44–46 Thus, the ability to alter Ig expression by VH–DH–JH recombination might be a mechanism for the diversification of immune responses other than somatic hypermutation.47 48 These findings suggest a remarkable genetic plasticity in subsets of antigen-reactive B lymphocytes in renal allograft.
In conclusion, our results provide evidence that there is more than one pathway to the onset and perpetuation of CD20 (+) B cell infiltration in acute rejection. In all patients but one with evidence of acute TCMR alone or associated with acute ABMR, CD20(+) B cells expressed highly mutated V genes without any signs of clonal expansion; it is unclear whether this pattern contributes to tissue destruction or (more probably) represents a non-pathological, unspecified bystander phenomenon.
In contrast, in the patient that had pathological evidence of glomerular inflammation, capillary thrombosis, and fibrinoid necrosis and diffuse C4d peritubular capillary and glomerular capillary deposition, we detected clonal expansion, suggesting a pressure for antigen and permanent local activation of the ectopic lymphatic tissue. This may well induce a more severe pattern of ABMR, through immune-mediated tissue damage.
Take-home messages
In all patients but one with evidence of acute TCMR alone or associated with acute ABMR, CD20(+) B cells expressed highly mutated V genes without any signs of clonal expansion.
This finding showed that Cd20(+) B cells become activated in peripheral lymphoid organs and subsequently migrate into renal tissue where they result in a local accumulation rather than a consequence of a local activation and differentiation probably resulting in a non-pathological, unspecified bystander phenomenon.
In the patient who had pathological evidence of glomerular inflammation, capillary thrombosis, fibrinoid necrosis and diffuse C4d peritubular capillary and glomerular capillary deposition, we detected clonal expansion, suggesting a pressure for antigen and permanent local activation of the ectopic lymphatic tissue. This may well induce a more severe pattern of ABMR, through immune-mediated tissue damage.
More than one pathway is involved to the onset and perpetuation of CD20(+) B cells infiltration in acute rejection.
References
Footnotes
Competing interests None.
Patient consent Obtained.
Ethics approval Ethics approval was provided by the Azienda Ospedaliera Universitaria Senese, University of Siena.
Provenance and peer review Not commissioned; externally peer reviewed.