Article Text

Abstract
Aims Infantile haemangioma is a tumour of the microvasculature characterised by aggressive angiogenesis during infancy and spontaneously gradual involution, often leaving a fibro-fatty residuum. The segmental distribution of a subgroup of infantile haemangioma, especially those associated with midline structural anomalies that constitute posterior fossa malformations–hemangiomas–arterial anomalies–cardiac defects–eye abnormalities–sternal cleft and supraumbilical raphe syndrome (PHACES), led us to investigate whether neural crest cells might be involved in the aetiology of this tumour.
Methods Immunohistochemical staining on paraffin embedded infantile haemangioma sections and immunocytochemical staining on cells derived from proliferating haemangioma cultures were performed.
Results The endothelium of proliferating infantile haemangioma contains abundant cells that express the neurotrophin receptor (p75), a cell surface marker that identifies neural crest cells, and also for brachyury, a transcription factor expressed in cells of the primitive mesoderm. The endothelium is also immunoreactive for the haematopoietic stem cell marker, CD133; the endothelial-haematopoietic stem/progenitor marker, CD34; the endothelial cell markers, CD31 and VEGFR-2; and the mesenchymal stem cell markers, CD29 and vimentin. Additionally, immunoreactivity for the transcription factors, Sox 9 and Sox 10, that are expressed by prospective neural crest cells was also observed. Cells from microvessel-like structures were isolated from in vitro cultured haemangioma tissue explants embedded in a fibrin matrix. Immunostaining of these cells showed that they retained expression of the same lineage-specific markers that are detected on the paraffin embedded tissue sections.
Conclusions These data infer that infantile haemangioma is derived from primitive mesoderm and that the cells within the lesion have a neural crest stem cell phenotype, and they express proteins associated with haematopoietic, endothelial, neural crest and mesenchymal lineages. The authors propose a model to account for the natural progression of infantile haemangioma based upon the multipotent expression profile of the primitive mesoderm and their neural crest stem cell phenotype to form all the cell lineages detected during infantile haemangioma proliferation and involution.
- Infantile haemangioma
- neural crest
- stem cells
- primitive mesoderm
- brachyury
- neurotrophin
- haemangioblast
- mesenchymal
- angiogenesis
- biological sciences
- cell biology
- haematopoesis
- vascular disease
Statistics from Altmetric.com
- Infantile haemangioma
- neural crest
- stem cells
- primitive mesoderm
- brachyury
- neurotrophin
- haemangioblast
- mesenchymal
- angiogenesis
- biological sciences
- cell biology
- haematopoesis
- vascular disease
Introduction
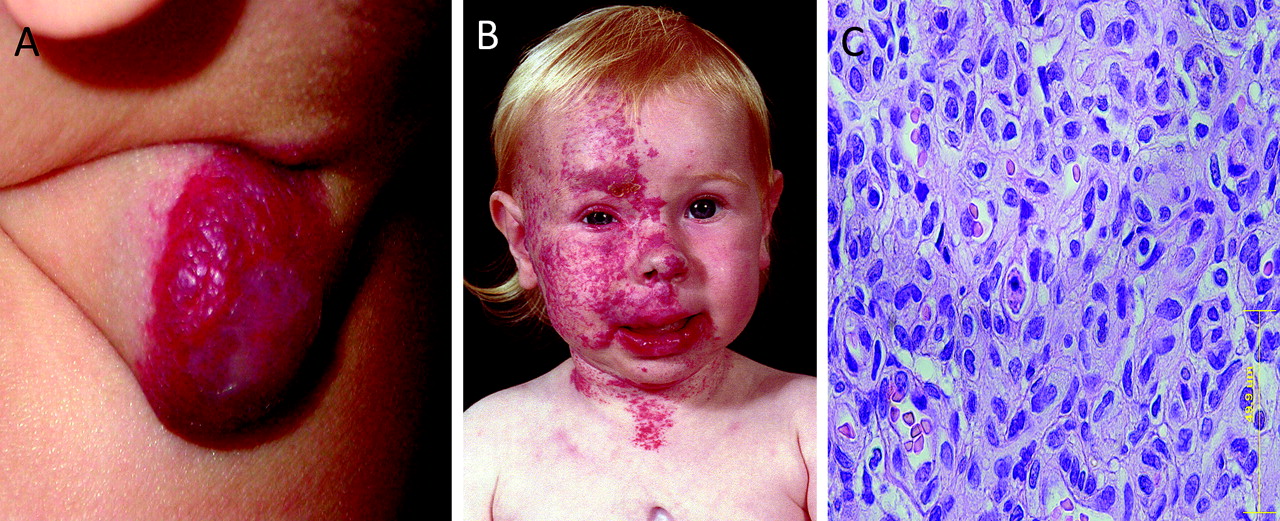
Infantile haemangioma (figure 1A) is the most common tumour of infancy affecting about 10% of white children, with a 3:1 female:male ratio and a preponderance in premature infants.1–3 It is typified by an initial rapid growth with aggressive angiogenesis during infancy, followed by spontaneous slow involution from 1–5 years of age in which there is diminished cellularity and increasing fibro-fatty deposition. After about 5–10 years of age, involution is complete, and lesions consist predominantly of fibro-fatty tissue with a few mature capillaries.1–4

(A) Ulcerated localised proliferating infantile haemangioma on the neck of a 4.5-month-old infant. (B) Girl with segmental facial haemangioma associated with a sternal cleft (posterior fossa malformations–hemangiomas–arterial anomalies–cardiac defects–eye abnormalities–sternal cleft and supraumbilical raphe syndrome (PHACES)). (C) H&E staining of a proliferating infantile haemangioma showing immature capillaries with tiny lumens lined by plump endothelial cells with an outer concentric pericyte layer.
The observation of the segmental distribution of a subgroup of infantile haemangioma,5 especially those associated with midline structural anomalies that constitute posterior fossa malformations–hemangiomas–arterial anomalies–cardiac defects–eye abnormalities–sternal cleft and supraumbilical raphe (PHACES) syndrome6 (figure 1B), has led us to investigate the role of neural crest cells in the aetiology of infantile haemangioma.
Infantile haemangioma consists of a heterogeneous mixture of cells including immature endothelial cells,7 myeloid haematopoietic cells8 such as mast cells,9 haematopoietic stem cells expressing CD13310 and mesenchymal stem cells.11 Recent reports suggest that the mesenchymal stem cells in haemangioma are recruited from the bone marrow and/or from the adjacent mesenchymal stem cell niches, and that these mesenchymal stem cells terminally differentiate into adipocytes as the lesion involutes.11 There is also growing evidence demonstrating that the stem cells in infantile haemangioma are derived from a mesodermal/mesenchymal phenotype.10 The identification of markers associated with endothelial and haematopoietic lineages in these same cells has also been reported.10
In this study we used immunofluorescence to investigate the expression of cell surface and cytosolic proteins associated with cells of the neural crest and primitive mesoderm.
Materials and methods
Immunohistochemistry
Immunohistochemical (IHC) staining was performed on 4 μm paraffin-embedded sections of localised infantile haemangioma from 12 patients, aged 3–10 (mean 6) months, according to a protocol approved by the Wellington Regional Ethics Committee.
Routine rehydration followed by antigen retrieval with boiling 10 mM sodium citrate (Sigma-Aldrich, St Louis, Missouri) was performed, and sections were washed in Tris-buffered saline (TBS; 20 mM Tris-HCl buffer, pH 7.6, containing 135 mM NaCl) containing 0.1% Tween 20 (Sigma-Aldrich). Autofluorescence was quenched with 0.5% sodium borohydride (Sigma-Aldrich) prepared in TBS, and the slides then blocked with 5% bovine serum albumin (Sigma-Aldrich) prepared in TBS containing 0.1% Tween 20 for 1 h. Primary antibodies were incubated overnight at 4°C. Primary antibodies used were CD34, 1:300; smooth muscle actin (SMA), 1:400; VEGFR-2, 1:300; CD133, 1:300; CD31, 1:400; p75, 1:200; Sox-9, 1:300; Sox-10, 1:300; CD29, 1:300; vimentin, 1:400; nestin, 1:200. Primary antibodies were obtained from Abcam (Cambridge, Massachusetts). Bound antibodies were visualised with the appropriate species-specific fluorochrome conjugated secondary antibody (goat antimouse Alexaflor-488 or chicken antirabbit Alexafluor-594 (Invitrogen, Auckland, New Zealand)), with the exception of VEGFR-2, which was detected with an antirabbit digoxigenin (Roche diagnostics, Auckland, New Zealand) conjugate followed by an antidigoxigenin-Rhodamine (Chemicon, Sydney, Australia). All slides were mounted in Antifade Gold (Invitrogen) containing DAPI as counterstain prior to visualisation.
Paraffin sections of uterine fibroid were used as appropriate negative controls to confirm the specificity of staining for p75, Sox 9, Sox 10 and brachyury. Control infantile haemangioma samples in which the primary antibodies were omitted showed minimal staining.
In vitro culture of tissue explants
Surgically resected proliferating infantile haemangioma (figure 1A) biopsies from three patients, aged 3, 6 and 8 months, were initially disinfected then cultured in a fibrin gel matrix as previously described by Tan et al12 in 24-well plates (Raylab, Auckland, New Zealand). The growth media were changed every 3 days, and following 3 weeks in culture, the cells emanating from the haemangioma explants were harvested from the fibrin gel using 50 caseinolytic units/well of Dispase (Becton-Dickson, Auckland, New Zealand). Dispase activity was quenched with dilution in MCDB (Sigma-Aldrich) media containing 5% Penstrep (Invitrogen), 0.5 μg/ml Amphotericin B (Sigma-Aldrich) and 5% foetal calf serum (Invitrogen) at a dilution of 1:10. Viable cells were then plated and allowed to adhere onto microscope slides divided into eight chambers (Becton-Dickson) at a density of 1×104 cells/chamber prior to fixation with ice-cold acetone/methanol (50:50) for 30 s.
Immunocytochemistry
Routine immunocytochemistry was performed using CD34, p75, CD29, VEGFR-2 & CD31. Staining of HeLa cells and the neuroblastoma cell line SY5Y were used as controls.
Microscopy
Staining was visualised and images captured using a Leica TCS 4D confocal laser-scanning microscope fitted with a krypton/argon laser (Leica Lasertechnik, Heidelberg, Germany).
Results
Proliferating infantile haemangima endothelium expressed primitive markers of multiple lineages
Proliferating infantile haemangioma consisted of immature capillaries with tiny lumens lined by plump endothelial cells with an outer concentric pericyte layer (figure 1C). These capillaries displayed an inner endothelium previously characterised by the endothelial marker CD311 and the haematopoetic-endothelial marker CD34,13 the outer pericyte layer confirmed by immunoreactivity (IR) to either smooth muscle actin (SMA)1 13 or nestin.14 Figure 2A shows IR for CD34 (green) in the endothelium and for SMA (red) in the pericyte layer. It is interesting to note that the IR for CD34 is predominantly localised to the luminal side of the endothelium. The endothelium characterised by IR for CD34 (green) in figure 2A,B also revealed IR for the endothelial cell marker, VEGFR-2 (figure 2B, red) and CD31 (figure 2C, green) as expected. Figure 2C shows that IR for CD133 (red) is localised to the CD31-positive cells. These cells that line the lumen also expressed the mesenchymal stem cell markers; vimentin (figure 2D, green), whereas the pericyte layer was confirmed by IR for SMA (figure 2D, red); and CD29 (figure 2E, red), with the pericyte layer confirmed by IR for nestin (figure 2E, green). The same luminal cells also showed IR for proteins associated with stem cells of primitive origin. The luminal cells demonstrated IR for the neural crest cell surface marker, p75 (figure 2F, red), distinct from the outer pericyte layer, showing IR to nestin (figure 2F, green). The endothelium staining positively for CD31 (figure 2G, green) was also IR to the primitive mesoderm transcription factor, brachyury (figure 2G, red). Consistent with this pattern of staining for primitive markers, the sections also showed IR for the neural crest cell associated transcription factors, Sox 9 (figure 2H, red) and Sox 10 (figure 2I, red). Figure 2, panels H and I, also show staining for CD34 (green), identifying the immature endothelial cells lining the capillary lumen.

Immunohistochemical staining of representative proliferating haemangioma sections from 12 patients. (A–I) All images are counterstained with DAPI (blue). Staining are as follows: (A) CD34 (green), SMA (red); (B) CD34 (green), VEGFR-2 (red); (C) CD31 (green), CD133 (red); (D) vimentin (green), SMA (red); (E) CD29, (red), nestin (green); (F) p75 (red), nestin (green); (G) CD31 (green), brachyury (red); (H) CD34 (green) Sox 9 (red); (I) CD34 (green), Sox 10 (red). Image 2A was taken at ×400, whereas images (B)–(I) were taken at ×200.
All infantile haemangioma tissue sections showed IR for Glut-1 (data not shown), the IHC marker for infantile haemangioma.2 Staining of negative controls, processed and analysed identically, was minimal (data not shown).
Cells emanating from proliferating infantile haemangioma explants expressed neural crest, endothelial, haemotopoietic-endothelial and mesenchymal cell marker
To determine whether the cells forming the microvessels that emanated from proliferating infantile haemangioma explants also expressed markers of multiple lineages, IR for the same array of markers used for IHC staining of tissue sections was evaluated on cells recovered from the fibrin matrix used to culture explanted tissue. Figure 3A shows staining for the neural crest cell surface marker, p75 (red), and the endothelial marker, CD31 (green). Doubly labelled cells appear orange in the presented overlayed image. The staining shown in figure 3A is in keeping with the p75 staining presented in figure 2F and the CD31 staining shown in figure 2C,G. The cells of the microvessels also co-expressed p75 (red) and CD34 (figure 3B, green). Staining for CD31 (figures 3C,D, green) in conjunction with VEGFR-2 (figure 3C, red) and CD29 (figure 3D, red) showed that the cells were positive for all these markers.

Immunocytochemical staining of representative microvessel-derived cells from three proliferating haemangioma biopsies. All images are counterstained with DAPI (blue). (A) CD31 (green), p75 (red); (B) CD34 (green), p75 (red); (C) CD31 (green), VEGFR-2 (red); (D) CD31 (green), CD29 (red).
Staining of HeLa cells and the SY5Y cells was performed under identical conditions to confirm the specificity of staining. HeLa cells were IR for vimentin, as expected, but showed minimal IR for the other antibodies used in this study (data not shown). SY5Y cells showed IR for p75 (data not shown).
Discussion
Our IHC data confirm previously documented staining patterns of proliferating infantile haemangioma in that IR for endothelial (VEGFR-215 and CD3116), haematopoietic-endothelial (CD3413) and the haematopoietic stem cell (CD13317) markers was found. In addition, our IHC data demonstrate the expression of proteins associated with neural crest cells within proliferating infantile haemangioma. IR for neural crest surface marker, p75,18 also known as neurotrophin receptor, and transcription factors, Sox 9 and Sox 10, is observed in the cells lining the immature capillaries of proliferating lesions. Sox 9 and Sox 10 belong to the group E Sox genes, which are expressed in prospective neural crest cells.19 Sox 9 transcription factor has been implicated in neural crest cell induction,19 whereas Sox 10 expression has been demonstrated in neural crest stem cells.20 21 The expression of Sox 10 transcription factor has been implicated in the maintenance of neural crest stem cell properties.20
Taken together, these data suggest a neural crest aetiology for infantile haemangioma. To our knowledge, this is the first report demonstrating the presence of cells expressing p75, Sox 9 and Sox 10 in infantile haemangioma.
The expression pattern of endothelial markers on the immature vessels of proliferating infantile haemangioma has been previously documented and is thought to be due to the angioblastic nature of these immature endothelial cells.7 Similar variable expression of endothelial markers has been shown in putative endothelial precursor cells,22 embryonic stem cell derived endothelial cells23 and angioblasts,24 25 confirming the primitive nature of these phenotypic endothelial cells.
Within proliferating infantile haemangiomas, we also observed coexpression of CD29 and vimentin on the cells lining the immature vessels. We infer that dual expression of CD29 and vimentin identifies a mesenchymal stem cell phenotype, consistent with the ability of haemangioma-derived mesenchymal stem cells to differentiate and form adipocytes in vitro, suggesting that the fibro-fatty tissue seen in involuted infantile haemangioma is derived from these mesenchymal stem cells. Other studies have reported that neural crest cells express CD2921 and vimentin.26 Cells staining for mesenchymal markers in proliferating infantile haemangioma have been isolated and shown to differentiate into mesenchymal-derived tissues, such as bone and fat,11 as well as neuro-glial cells.10 These observations, combined with the demonstration of neural crest stem cell markers on the luminal surface of the immature capillaries that we report, infer that these cells are embryonic stem cell-derived neural crest stem cells with the ability to form cells of both mesenchymal and peripheral neuronal lineages as previously described in the literature.27
Expression of CD34, a marker of haematopoietic-endothelial cells,28 and CD133, a marker of haematopoietic stem/progenitor cells,29 suggests a primitive haematopoietic phenotype. The coexpression of CD133, CD34 and VEGFR-2 is characteristic of stem cells and progenitor cells that have the ability to differentiate into mature endothelial cells.30 31 Haemangioblasts have been suggested as the common precursor of both haematopoietic and endothelial cells32 through a haemogenic endothelium stage33 34 and have been proposed to express CD34 and VEGFR-2.30 35 We have recently demonstrated the expression of haematopoietic markers on the phenotypic endothelial cells of the immature capillaries of proliferating infantile haemangioma lesions, and we infer that the immature capillaries are structural haemogenic endothelium.36 The expression of brachyury by these same cells indicates that they are derived from the primitive mesoderm and have the ability to form haemangioblasts.37 Foetal mesenchymal stem cells, on the other hand, are dually positive for vimentin and CD29 but are negative for both haematopoietic and endothelial markers.38 Early embryonic vasculogenesis, however, involves the formation of angioblasts from mesodermal cells,24 and the plasticity of mesenchymal cells to differentiate into endothelial cells requires various growth factors, among which, VEGF plays a prominent role.24 39 These data show that the endothelium of proliferating infantile haemangioma expresses proteins associated with the primitive mesoderm and neural crest stem cells, as well as the previously reported markers of haematopoietic, endothelial and mesenchymal stem cell lineages. It is interesting to speculate that the expression of markers for all four lineages that we report infers that infantile haemangioma has a primitive mesodermal origin with a neural crest stem cell-like phenotype, and that this primitive mesodermal neural crest phenotype has the ability to potentially differentiate down all four lineages, but that is beyond the scope of this paper.
The segmental migratory patterns of neural crest cells in early embryogenesis40 may explain the segmental distribution of infantile haemangioma including those occurring in PHACES syndrome6. This distribution suggests that infantile haemangioma is a developmental disorder of aberrant neural crest stem cells. These cells we propose have the ability to give rise to both the mature endothelial1 and the mature mesenchymal derivatives11 observed within involuted infantile haemangioma. We suggest that the fibro-fatty tissue characteristic of involuted infantile haemangioma is due to preferential differentiation of these neural crest cells towards the mesenchymal lineage and apoptosis of the endothelial component, possibly in response to the low levels of VEGF1 and/or other pro-angiogenic cytokines. Whether these neural crest stem cells give rise to intermediary cells such as haemangioblasts and the mesenchymal stem cells, or whether they have the ability to differentiate directly to form cells of all four lineages is the topic of further investigation.
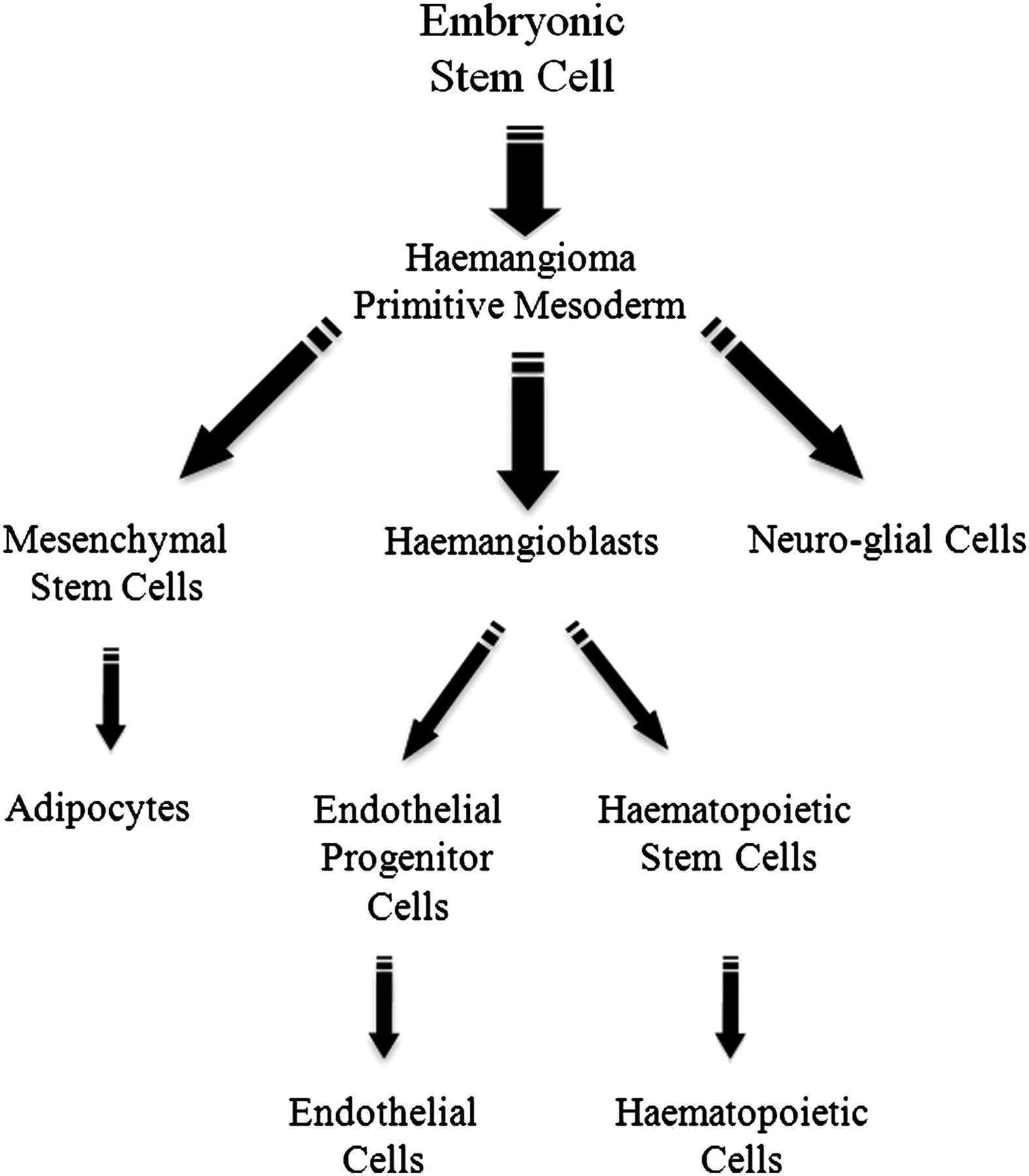
Figure 4 shows a possible model to account for all the cell types observed in infantile haemangioma that is consistent with, and may be able to account for, the biological characteristics of infantile haemangioma. In this model, we propose that an embryonic stem cell gives rise to infantile haemangioma-initiating primitive mesodermal neural crest cells that proliferate and give rise to haemangioblasts, mesenchymal stem cells and neuro-glial cells. During early tumour growth, haemangioblasts and their progeny predominate, such that the lesion expresses abundant endothelial and haematopoeitic markers. As infantile haemangioma involutes, and levels of VEGF decrease, the endothelial component is reduced, and the mesenchymal component predominates. The mesenchymal stem cells eventually terminally differentiate to form the adipocytes which make up the fibro-fatty deposits within involuted lesions.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Model illustrating the origin of the cell lineages observed in the progression of infantile haemangioma. Embryonic stem cells are proposed to give rise to the haemangioma primitive mesodermal neural crest stem cells. These cells are able to form mesenchymal stem cells, haemangioblasts and neuro-glial cells. During the proliferative phase of haemangioma, haemangioblasts give rise to the immature endothelial cells that predominate the lesion. Haemangioblasts generate cells of haematopoietic lineage such as mast cells that become highly prominent during the involuting phase of infantile haemangioma. Later in the involuting and involuted phases, the neural crest stem cells give rise mainly to mesenchymal stem cells that are able to differentiate to form the adipocytes that make up the fibro-fatty tissue in the involuted lesion.
Take-home message
Infantile haemangioma endothelium is derived from primitive mesoderm with a neural crest stem cell phenotype, and they co-express proteins associated with haematopoietic, endothelial and mesenchymal lineages.
References
Footnotes
Equal senior authors: STT, DJD.
This paper was presented, in part, at the Australian and New Zealand Vascular Anomalies Interest Group Meeting, 22 October 2009, Melbourne, Australia; the New Zealand Association of Plastic Surgeons' Annual Scientific Meeting, 21 November, 2009, Auckland, New Zealand; the Royal Australasian College of Surgeons' 46th Surgical Research Society Meeting, Adelaide, Australia, 20 November 2009; and the International Society for the Study of Vascular Anomalies Workshop, Brussels, Belgium, 21–24 April 2010.
Funding We wish to thank the Wellington Regional Plastic Surgery Unit Research & Education Trust; the Wellington Medical Research Foundation, the Surgical Research Trust; and Pub Charity for their financial support of this project. TI is supported by a Royal Australasian College of Surgeons' Foundation for Surgery Scholarship.
Competing interests None.
Patient consent Obtained from the parents.
Ethics approval Ethics approval was provided by the Wellington Regional Ethics Committee.
Provenance and peer review Not commissioned; externally peer reviewed.