Article Text

Statistics from Altmetric.com
Introduction
Colorectal cancer is one of the commonest malignant neoplasm worldwide, and over 35 000 colorectal cancers were diagnosed in England in 2004.1 Unfortunately, over 57% have regional or distant spread of their disease at the time of diagnosis.2 More recently, research has focused on molecular biomarkers to indicate progression of a disease or the susceptibility of the disease to a given treatment.3
The CCN family comprises cysteine rich-61 (Cyr61/CCN1), connective tissue growth factor (CTGF/CCN2) and nephroblastoma overexpressed (Nov/CCN3). These proteins stimulate mitosis, adhesion, apoptosis, extracellular matrix production, growth arrest and migration of multiple cell types.4 The role of the CCN members in cancer is far from clear, and the results in the literature remain controversial.
To date, the expression of CTGF and Cyr61 mRNA and protein has not been studied together in colorectal cancer specimens. In this study, we determined the in vitro effects of hypoxia on Cyr61 and CTGF expression in colorectal cancer cell lines. We assessed the expression of Cyr61 and CTGF mRNA and protein in colorectal cancer. This was compared with various clinicopathological parameters.
Materials and methods
Ethical permission was sought to allow the use of patient tissue samples for the purpose of this study that included 39 patients who had a primary colorectal tumour (table 1).
Clinicopathological characteristics of the study population
Cell culture
HT29 and Caco-2 cells derived from a human colorectal cancer were cultured in McCoy's 5A Medium (Invitrogen, Carlsbad, California) supplemented with 10% fetal bovine serum and incubated in 5% carbon dioxide. To induce hypoxia, cells were treated with 100 μM of desferrioxamine (Sigma, St Louis, Missouri). Caco-2 cells were treated with desferrioxamine for 24 and 48 h. Normoxic cell lines were cultured in the same way without the use of desferioxamine.
RNA isolation and RT-PCR
Isolation of mRNA and reverse transcription was carried according to the manufacturer's instructions.5 Briefly, a 10 μl aliquot of mRNA was reverse-transcribed into single-stranded cDNA using Avian Myeloblastosis Virus Reverse Transcriptase (AMV-RT) and random hexamers (Promega, Madison, Wisconsin). Samples were subsequently incubated at 42°C for 60 min. The cDNA (1 μl) was amplified with the forward (F) and reverse (R) primers by PCR. Thermal cycling conditions were as follows: one cycle of 98°C for 3 min, 60°C for 3 min and 72°C for 1 min and 40 cycles of 94°C for 30 s, 60°C for 30 s and 72°C for 30 s. At the initial annealing phase, thermostable Taq DNA polymerase (Promega) was added to each PCR reaction. PCR-amplification products were loaded onto a 3% agarose gel containing 50 μg/100 ml ethidium bromide. The primers used for RT-PCR are outlined in table 2.
Primer sequences used for this study
Real-time quantitative PCR
Briefly 4 μl of cDNA was amplified in a 6 μl reaction solution containing, 5 μl of SYBR Green Jumpstart Taq Ready Mix, 0.2 μl of both the forward and reverse primer and 0.6 μl of sterile H2O. Thermal cycling conditions were as follows: one cycle of 98°C for 3 min and 60°C for 5 min and 40 cycles of 94°C for 15 s and 60°C for 30 s. Comparative gene expression was calculated using the Pfaffl equation.6
RNA extraction of paraffin-embedded tissues
The use of formalin-fixed tissues as a source of RNA requires appropriate RNA extraction and amplification methods.7 RNA was extracted from paraffin-embedded tissues as follows, using 500 μl 0.1% sodium dodecyl sulfate (pH 7.3) and Tris/HCl (pH 8.0). This was then kept overnight in 5 μl of proteinase K (Sigma) (1 μg/ml) at 65°C. Total RNA was extracted using TRIzol reagent (Invitrogen) according to the manufacturer's protocol.
A universal reverse transcriptase primer (URTP) was designed that targets the 3′ poly (A) tail of mRNA and adds a unique tag sequence not otherwise found in the human genome. The RNA (10 μl) was added to 1.5 μl of CTGF URTP, 1.5 μl of 18S URTP and 2 μl of sterile water. This was heated to 70°C for 5 min and then reverse-transcribed as stated before.
Real-time PCR was subsequently performed using a specific forward (F) primer and a universal reverse primer that binds to the universal reverse transcriptase tag primer (table 2). This priming method has the advantage of being sensitive and specific.8
Immunohistochemistry
For immunostaining, tissue sections were subjected to a heat-mediated antigen retrieval procedure using 0.01 M citrate buffer. Polyclonal antibody preparations specific for CTGF (ab-6992; Abcam, Cambridge, UK) and CYR61 (sc-13100; Santa Cruz Biotechnologies, Santa Cruz, California) were used.9 10 CTGF antibody was applied to the slides at a dilution of 1:1000, while Cyr61 antibody was used at a dilution of 1:2000. A secondary biotinylated swine antirabbit antibody (Dako, Carpintería, California) was used at a dilution of 1:800 with strep–avidin–biotin alkaline phosphatase (Dako) complex. Slides were stained with fast red substrate.
Cyr61 immunostaining was determined by the H score calculated for both the tumour cell and extracellular matrix expression combined.11 A score was devised to assign absent, weak, moderate and strong staining intensity to CTGF.
Statistical analysis
Statistical analysis of cell lines using qRT-PCR was based on Student t tests. Comparisons between groups were made by a one-way analysis of variance (ANOVA) using Bonferroni corrections. Differences between immunohistochemistry data were detected using Mann–Whitney and Kruskal–Wallis tests. A trend analysis was performed using the Jonckheere–Terpstra test.
Results
In vitro effects of hypoxia on mRNA expression of Cyr61 and CTGF in colorectal cancer cell lines
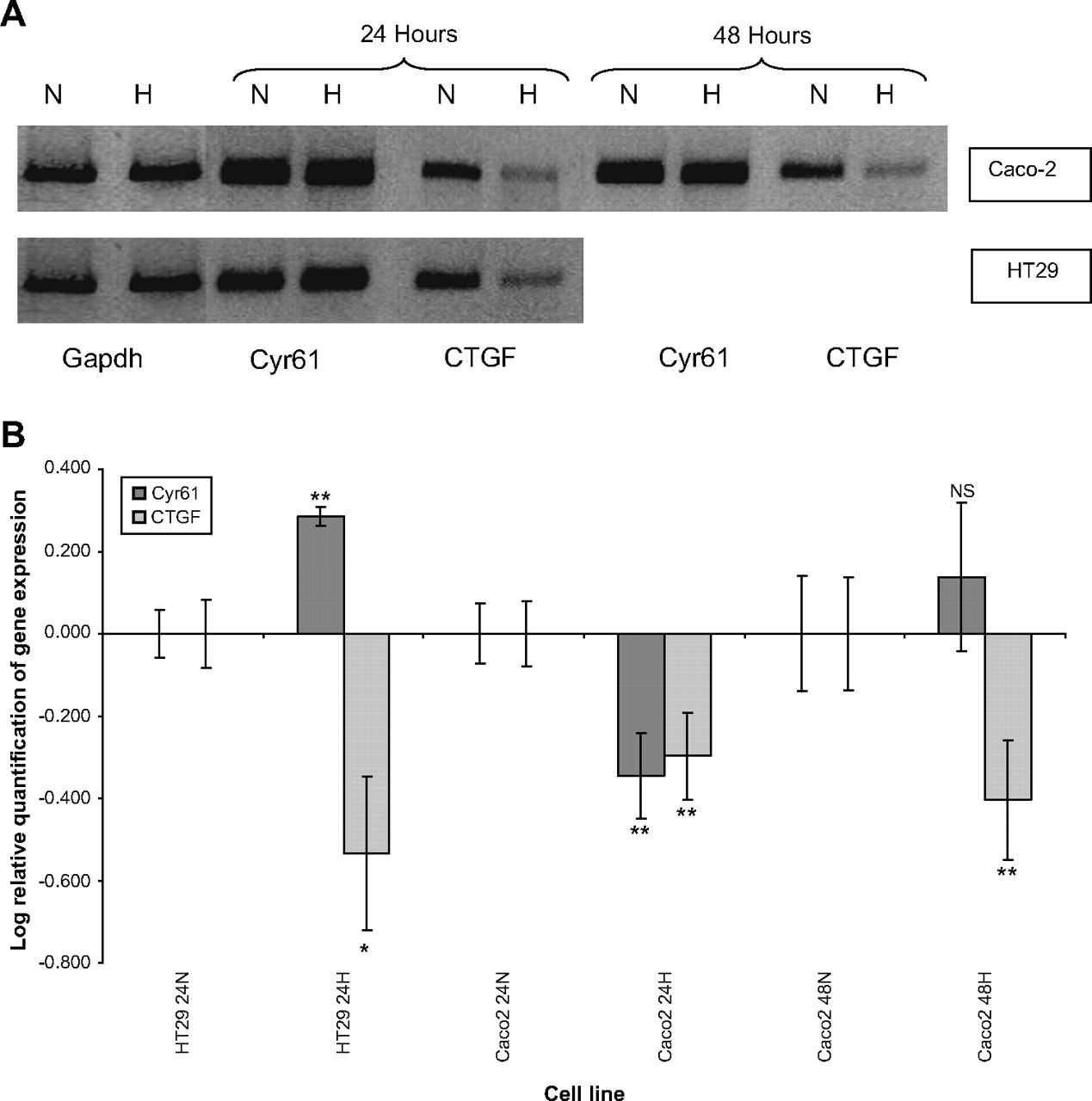
CTGF mRNA expression consistently decreased in HT29 and Caco-2 cell lines when exposed to desferrioxamine (p<0.01). CTGF was downregulated at both 24 and 48 h induction of hypoxia in Caco-2 cell lines. Further downregulation of CTGF was observed at 48 h compared with 24 h. Cyr61 was upregulated in the HT29 cell line (p<0.01) but downregulated in the Caco-2 cell line (p<0.01). Cyr61 mRNA was not significantly different from normoxic conditions at 48 h of hypoxia in the Caco-2 cell line (figure 1).
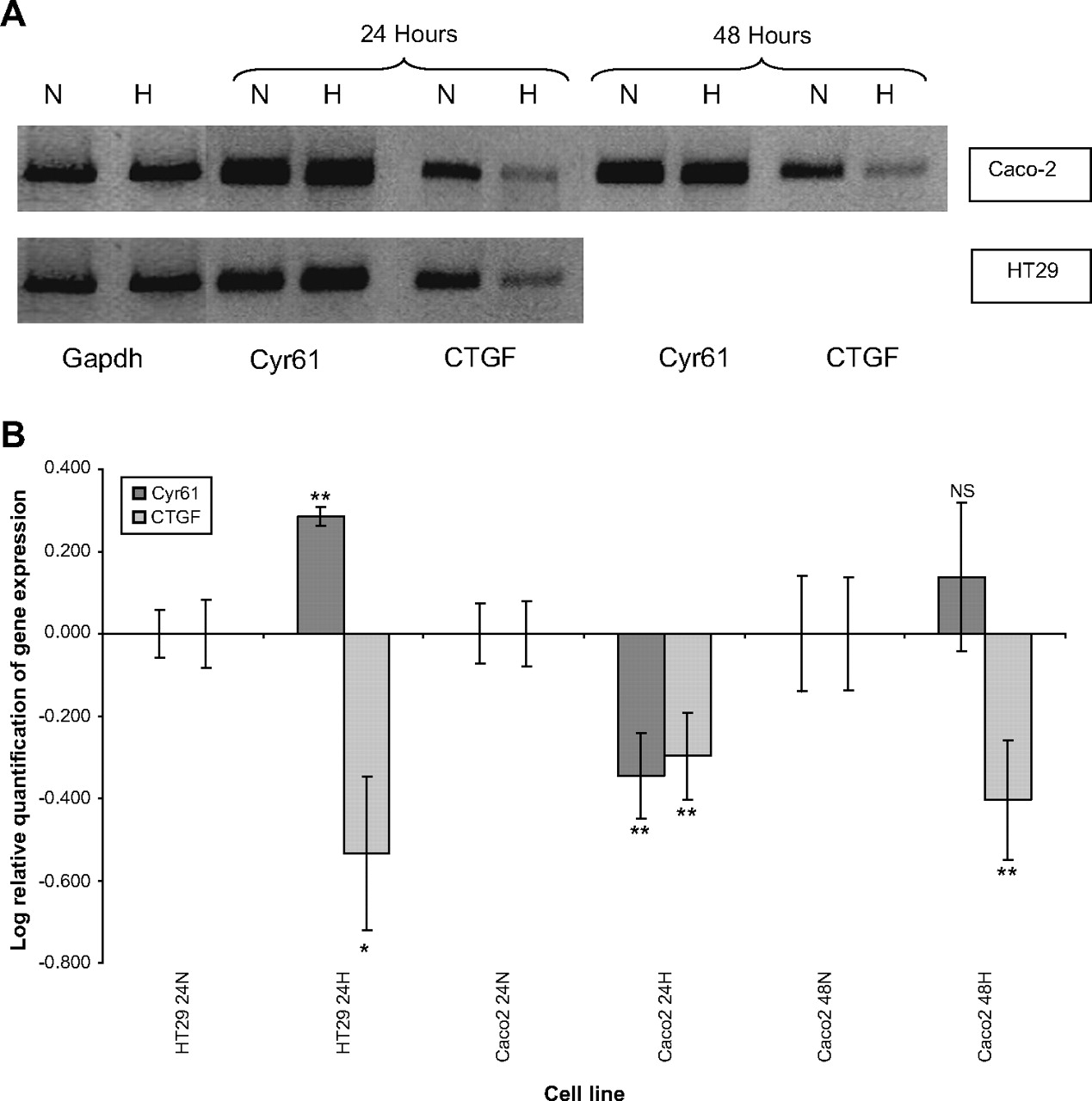
In vitro analysis of cysteine rich-61 (Cyr61) and connective tissue growth factor (CTGF) mRNA in colorectal cancer cell lines exposed to hypoxia at 24 and 48 h. (A) RT-PCR analysis of Cyr61 and CTGF expression in HT29 and Caco-2 colorectal cancer cell lines in normoxic and hypoxic (H) conditions. Control cultures were kept under normoxia (N). Glyceraldehyde-3-phosphate dehydrogenase bands are shown to confirm equal loading of RNA. (B) Log relative quantification of Cyr61 and CTGF in the HT29 and Caco-2 colorectal cancer cell lines using desferrioxamine-induced hypoxia (H) referenced to each cell line's normoxic (N) control by qRT-PCR (40 cycles). The Caco-2 cell lines were exposed to desferrioxamine-induced hypoxia for 24 and 48 h. Error bars represent the SE of the results due to variation in triplicate results. *p<0.05; **p<0.01.
Expression of CTGF and Cyr61 mRNA in colorectal cancer tissues
CTGF and Cyr61 mRNA were all readily detectable in all tumour tissue samples and control mucosa specimens (figure 2). CTGF and Cyr61 mRNA was expressed to a greater degree in the majority of colorectal cancer specimens (table 3) compared with normal colon tissue (p<0.01). This was particular evident in Dukes' stage A and B tumours. There was no significant difference between CTGF and Cyr61 mRNA expression between Dukes' stage C tumours and normal colon tissue. However, there was no statistically significant difference in CTGF and Cyr61 mRNA with advancing Dukes' stage (figure 3).

Log relative quantification (log RQ) of cysteine rich-61 (Cyr61) and connective tissue growth factor (CTGF) mRNA expression in colorectal cancer paraffin embedded tissues (N=28) and histologically normal bowel from patients with colorectal cancer (N=7) by qRT-PCR. Cases (1–28) are referenced to N2.
Association of cysteine rich-61 (Cyr61) and connective tissue growth factor (CTGF) mRNA expression with clinicopathological parameters in colorectal cancer, as determined by qRT-PCR
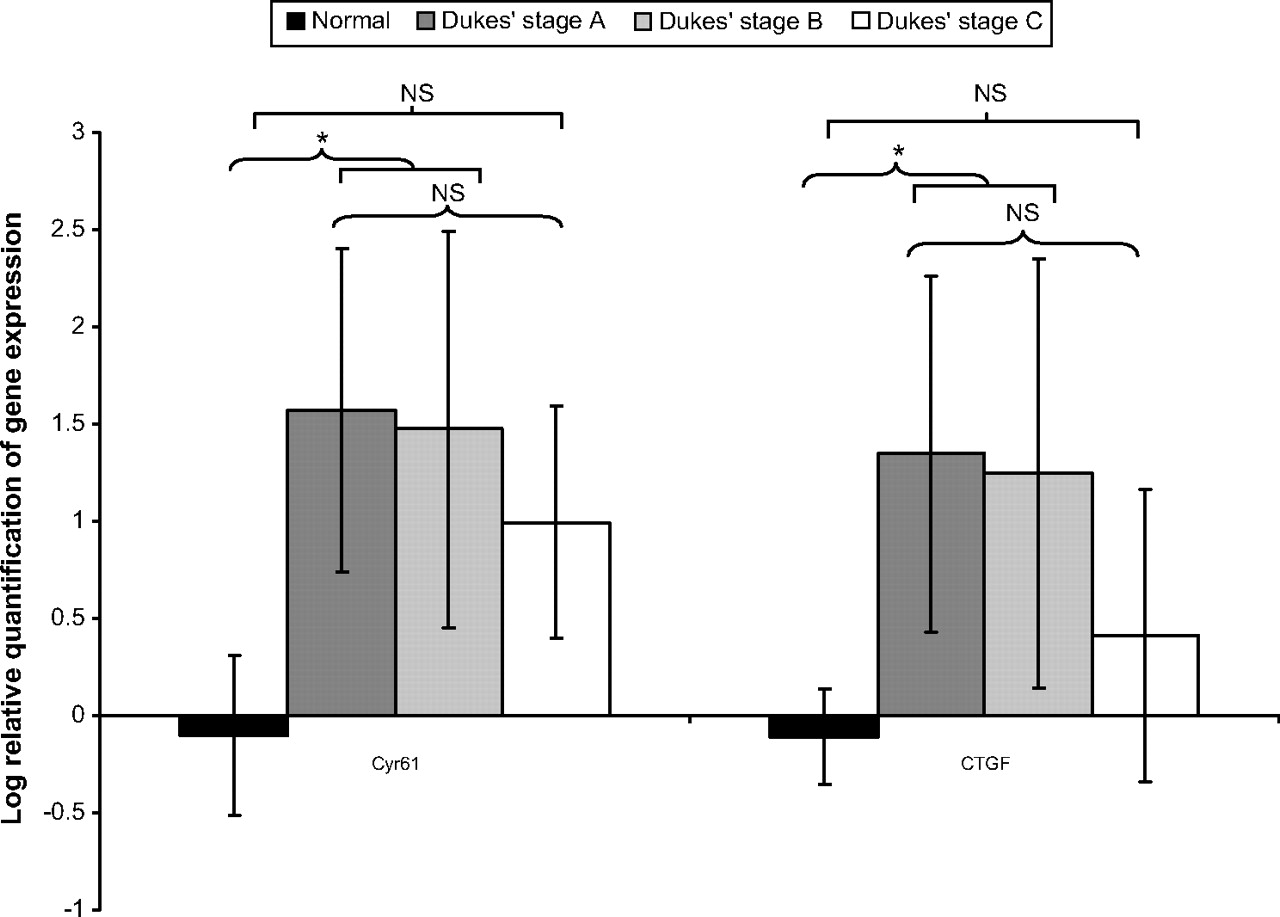
Log relative quantification of cysteine rich-61 (Cyr61) and connective tissue growth factor (CTGF) mRNA expression with Dukes' stage in colorectal cancer paraffin embedded tissues and normal colorectal tissues. *p<0.05. NS, not significant.
We did not identify any statistically significant difference between CTGF and Cyr61 mRNA expression with other pathological parameters such as TNM stage, tumour differentiation or extramural vascular invasion.
Expression of CTGF and Cyr61 protein levels in colorectal cancer tissues
Immunohistochemistry was performed on 39 cases for CTGF and 28 cases for Cyr61 (11 cases were omitted because of a lack of tissue).
CTGF protein expression was present in 95% of cases. Tumour tissues showed varying intensities of staining, with the most prominent staining in the cytoplasm of tumour cells (figure 4). There was an increase in the intensity of staining in advancing Dukes' stage, tumour stage and node stage (table 4). Protein expression was higher in Dukes' stage C compared with Dukes' stage A (p<0.01). There was no significant difference between Dukes' stages A or C tumours compared with Dukes' stage B tumours (figure 5A). Also, there was an increase (p<0.01) in CTGF protein expression in pT3 and pT4 staged tumours compared with pT1 and pT2 tumours (figure 5B). There was a significant increase (p<0.01) in expression of CTGF protein in tumours staged as pN1 and pN2 compared with pN0 (figure 5C).

Connective tissue growth factor immunostaining in colorectal cancer tissue showing diffuse cytoplasmic staining of different grades of intensity: (A) strong cytoplasmic staining; (B) moderate cytoplasmic staining; (C) limited staining of normal colorectal tissue; (D) control sections of colorectal cancer with no primary antibody. Cysteine rich-61 showed (E) prominent staining in the lamina propria of normal colorectal tissue and (F) strong tumour staining.
Association of connective tissue growth factor (CTGF) and cysteine rich-61 (Cyr61) protein levels with clinicopathological parameters in colorectal cancer cases

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Immunohistochemical expression of connective tissue growth factor (CTGF) protein levels with (A) Dukes' stage (B) tumour (T) stage and (C) Node (N) status in colorectal cancer paraffin embedded tissues. **p<0.01. NS, not significant. Error bars represent 95% CIs.
Cyr61 staining was present in 82% (23/28) of cases. Staining was present in both the tumour and extracellular matrix (figure 4). There were no significant differences between Cyr61 protein expression and clinicopathological parameters (table 4).
Discussion
In the present study, we have studied the expression of mRNA and protein expression in colorectal cancer using qRT-PCR and immunohistochemistry. We have shown for the first time an increased expression of CTGF and Cyr61 genes and protein in colorectal cancer. Expression of CTGF and Cyr61 has led to conflicting roles in a variety of cancers. Overexpression of CTGF, compared with normal control specimens, was observed in oesophageal cancer, pancreatic cancer and gliomas.12–14 In oesophageal adenocarcinoma and breast cancer, higher levels of CTGF are associated with more advanced disease.12 15 In contrast, higher levels of CTGF are associated with a better prognosis for lung adenocarcinoma and colorectal cancer.16 17
Similarly, compared with normal tissues, Cyr61 overexpression was observed in ovarian cancer, breast cancer, gliomas and benign prostate hyperplasia.14 18–20 In contrast, downregulation was observed in endometrial cancer, thyroid cancer and non-small-cell lung cancer.21–23 Furthermore, Cyr61 expression increased in more advanced cases of breast cancer and glioma, suggesting it may have a role in the progression of tumours.14 24 Of note, in our study, Cyr61 and CTGF gene expression was overexpression in primary cancers, but its expression was reduced to levels comparable with that of normal colorectal epithelium in advanced Dukes' stage. We did not find a clear link between the level of Cyr61 expression and colorectal cancer progression based on Dukes' stage or TNM stage. In another study, upregulation of Cyr61 expression was present in young patients with early-onset colorectal cancer.25 CTGF and Cyr61 could act as early tumour markers that can detect at-risk patients.
Variations in levels of Cyr61 and CTGF could be related to other factors within the tumour microenvironment. Studies have indicated that hypoxia may cause a more aggressive tumour, resistant to treatment.26 In this study, we induced hypoxia in colorectal cancer cell lines using desferrioxamine at 24 and 48 h. We showed that CTGF was downregulated in colorectal cancer cell lines when exposed to hypoxia (p<0.01), and further downregulation was noted at 48 h. Hypoxia-mediated downregulation of CTGF could lead to a more invasive tumour. Cyr61 expression varied with cell lines, suggesting a complex mechanism of control. In one study, downregulation of Cyr61 was thought to be associated with more aggressive melanoma cell lines.27
We found a correlation between increased protein levels of CTGF with advanced Dukes' stage and TNM stage. In contrast, increased CTGF protein expression in colorectal cancer was related to a better prognostic outcome in another study.28 CTGF immunostaining was present in the extracellular matrix but predominantly in the cytoplasm of tumour cells. This is similar to several other tumour types, including oesophageal and pancreatic tumours.12 13 We did not quantify CTGF protein staining within the extracellular matrix. The secretion of the peptide into the extracellular matrix could account for differences in this study compared with others and account for differences in the mRNA and protein quantification. One study showed exposure of hypoxia on trophoblasts affected mRNA transcript levels but had no effect on cellular protein levels and, furthermore, increased CTGF protein secretion.29
Understanding how CCN proteins function to either promote or inhibit tumorigenesis will require further investigation. Nevertheless, the strong upregulation of CCN expression in colorectal cancer has implicated their role in tumour development and tumour progression. The strong upregulation of these genes early in colorectal cancer suggests that CCN signalling pathways may be useful targets for novel anticancer therapy and could be used to identify colorectal tumours early on in disease progression.
Take-home messages
Connective tissue growth factor and cysteine rich-61 play a key role in tumorigenesis in several human malignancies.
The gene expression of these factors was overexpressed in colorectal cancer but reduced at the advanced Dukes' Stage. Hypoxia alters the expression of these genes in colorectal cancer cell lines.
We found a correlation between increased protein levels of CTGF with advanced Dukes' stage and TNM stage. The secretion of the peptide into the extracellular matrix could account for differences in the mRNA and protein quantification.
There was no correlation between Cyr61 and clinical outcomes.
References
Footnotes
Funding Jean Shanks Foundation.
Competing interests None.
Ethics approval Ethics approval was provided by the Leicestershire, Northamptonshire and Rutland Research Ethics Committee.
Provenance and peer review Not commissioned; externally peer reviewed.