Article Text

Abstract
Aims The pathogenesis of thyroid-associated orbitopathy (TAO) remains unclear. The aim of this study is to elucidate the gene expression profile of orbital fat from patients with active, but untreated, TAO.
Methods A case–control gene expression study was conducted using test samples of orbital fat from TAO patients and control orbital fat specimens; apart from drugs to control thyrotoxicosis, the TAO patients had received no treatment for orbital disease. cDNA expression analysis was performed using the Affymetrix GeneChip Human Genome U133 Plus 2.0 platform and validated using quantitative PCR.
Results The highest-ranked differentially expressed genes were dominated by IGF-1 signalling genes. These include IGF-1, IGF-1 receptor binding/signalling genes, such as SOCS3 and IRS2, and downstream signalling and transcriptional regulators, such as SGK (PDK/Akt signalling) and c-JUN. Our microarray data also demonstrate dysregulation of wingless-type MMTV (Wnt) signalling gene expression, including Wnt5a, sFRPs and DKK.
Conclusion Altered Wnt signalling confirms previous array findings. Further investigation of the role of Wnt signalling in TAO pathogenesis is warranted. These data also provide the first evidence of dysregulation of IGF-1 pathway genes in TAO tissue, further strengthening the evidence for the role of IGF-1 signalling in the pathogenesis and potential treatment of TAO.
- Graves orbitopathy
- IGF-1
- Wnt
- thyroid eye disease
- eye
- cell biology
- collagen
Statistics from Altmetric.com
Introduction
Thyroid-associated orbitopathy (TAO) is a common disorder affecting patients with autoimmune disease of the thyroid. The clinical manifestations of TAO include extraocular muscle inflammation and fibrosis, upper eyelid retraction and proptosis. These effects can cause significant morbidity, with an estimated 20–50% of patients with autoimmune thyrotoxicosis developing ophthalmic symptoms and 5% experiencing optic neuropathy that can lead to irreversible vision loss.1 ,2 Optic neuropathy results from an increased volume of soft tissues within the rigid bony orbit, which is the result of three distinct but related processes: (1) inflammatory swelling of the extraocular muscles; (2) deposition of hyaluronan (HA), a highly hydrophilic glycosaminoglycan whose structural and electrochemical properties increase the water content (and thereby volume) of the orbital fat compartment3 and (3) adipogenesis, the differentiation of orbital fibroblasts into adipocytes,4 with subsequent increased orbital fat content.
While a definitive unified model for the pathogenesis of TAO remains elusive, it is currently believed that orbital tissues are infiltrated by immunocompetent cells that release inflammatory mediators activating orbital fibroblasts. When activated, orbital fibroblasts produce proinflammatory cytokines such as interleukin 1a (IL-1a), IL-1b, CD154 and IL-6. These cytokines have been shown to activate genes such as prostaglandin H synthase (PGHS)-2, IL-6, IL-8, HA synthase and UDP-glucose 6-dehydrogenase (UGDH), and also genes regulating adipocyte proliferation.5 ,6 Mechanisms involving insulin-like growth factor-1 (IGF-1) have been strongly implicated in the pathogenesis of TAO. It has been demonstrated that orbital fibroblasts from patients with autoimmune thyrotoxicosis express high levels of IGF-1 receptor (IGF-1R)7 and stimulation of IGF-1R in these cells leads to an increased expression of HA, IL-16 and RANTES (regulated upon activation, normal T-cell expressed and secreted),7–9 possibly acting through an IGF-1R complex co-localising with the thyroid-stimulating hormone receptor (TSHR).10 Both B and T lymphocytes have also been found to express high levels of IGF-1R.11 ,12
The mechanisms of adipogenesis and HA production in TAO have been investigated largely using in vitro cultures of orbital fibroblasts which fail to account for the complex interactions of heterogeneous cell types mediating autoimmune stimulation in TAO. The identification of differentially expressed genes in orbital adipose tissue from patients with active disease offers the prospect of providing new insights into the molecular mechanisms underlying the pathogenesis of TAO. One approach to identifying dysregulated genes has been the use of synthetic oligonucleotide cDNA microarrays, which allow for the simultaneous evaluation of the expression of tens of thousands of genes.
There have been three previous expression microarray studies comparing active TAO with normal orbital tissue: Kumar et al13 identified upregulation of several adipocyte regulatory genes, as well as soluble frizzled-related protein 1, a wingless-type MMTV (Wnt) signalling gene. The potential involvement of Wnt signalling dysregulation was further confirmed in a study identifying downregulation of secreted frizzled-related protein 3 (sFRP3) in active TAO using PCR only.14 Lantz et al, in a study of five TAO patients and five controls, identified upregulation of adipocyte-related immediate early genes in orbital fat from TAO patients.15 Although of unclear significance, an upregulation of ceroid lipofuscinosis neuronal genes 2 and 3 (CLN2, CLN3) has also been identified in a small sample of two TAO and two control cases.16
The aim of this investigation is to use the current gold standard transcriptome-level microarray platform to identify differences in gene expression between control orbital fat from patients with untreated, active, inflamed TAO and that from controls without TAO or orbital inflammation.
Methods
Orbital tissue harvesting
This investigation was conducted in accordance with local and regional ethics approval.
Five orbital fat samples, which were taken for diagnostic purposes from patients with active TAO, were identified from the pathology library at the UCL Institute of Ophthalmology. Paraffin-embedded tissues were chosen, as it is no longer common practice to perform an orbital biopsy for active Graves' orbitopathy, and these samples were identified from a historical archive of tissue blocks. Tissue samples of orbital fat from untreated TAO patients are rare and only five samples were identified over the last 16 years from one of the largest international centres for orbital disease. Four control samples were harvested prospectively from patients undergoing removal of lateral orbital subconjunctival fat herniation. Control samples were taken from identical sites (lateral orbit) to disease tissue samples and also fixed and embedded in paraffin to ensure methodological consistency. The clinical characteristics of the patients from whom the diseased tissues were collected are summarised in table 1. All patients had active disease, with progressive proptosis as the main feature at the time of biopsy, and had not been on any immunomodulatory treatments.
Clinical characteristics of TAO group
RNA extraction and microarray gene expression profiling
RNA extraction was performed using a validated proprietary automated extraction technology that can undertake high quality arrays from stored formalin fixed paraffin embedded archival tissue (Response Genetics, Inc; Los Angeles, California, USA patent application 20090092979), as previously described.17 This system has been designed to optimise the yield of high molecular weight RNA fragments.
The GeneChip Human Genome U133 Plus 2.0 (Affymetrix, Santa Clara, California, USA) transcriptome-level cDNA microarray platform was used, which covers the expression of over 47 000 transcripts and variants, including 38 500 characterised human genes, and provides expression analysis for an extremely wide range of genes. cDNA was hybridised to the array chip following the standard Affymetrix protocols. Arrays were scanned on a GeneChip 3000 7G Scanner and data collected as ‘.DAT’ files before conversion to ‘.CEL’ files for analysis. For quantitative validation of mRNA expression, quantitative reverse transcriptase PCR was performed using the TaqMan Gene Expression Assay (Applied Biosystems, Carlsbad, California, USA) following the manufacturer's instructions. GAPDH was used as an endogenous control for normalisation.
Gene array ‘*.CEL’ files were analysed using Genespring GX V.10.01 2100 (Agilent, Santa Clara, California, USA). Gene-level expression values were generated using ‘Robust multiarray average’, which is a quantile-based normalisation tool for normalising microarray data.18 Normalised expression data for the two groups were compared using analysis of variance and an α-risk of 0.05 was taken as being statistically significant. For quantitative PCR validation, analysis was performed using GraphPad Prism software and groups were compared using the Student t test.
Results
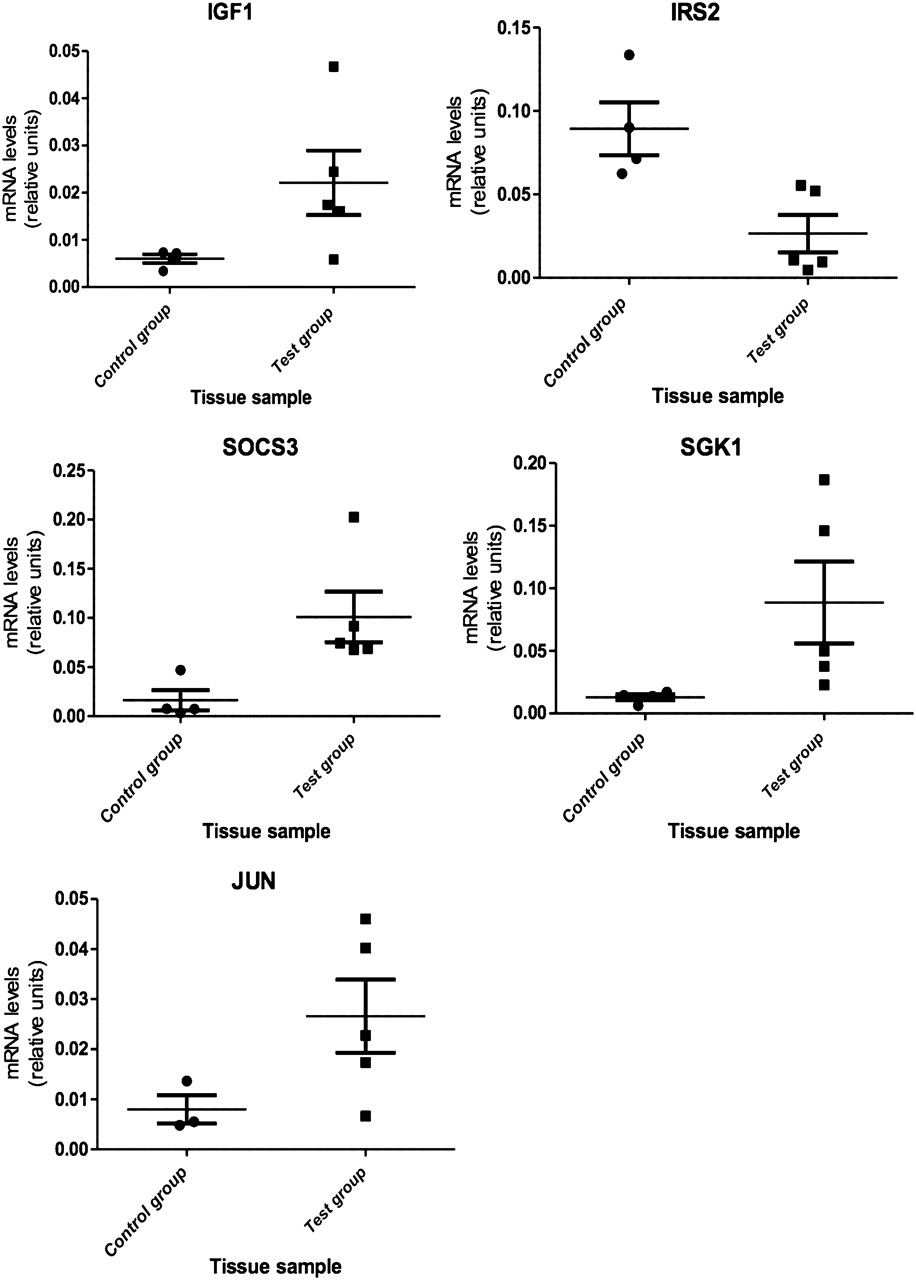
Microarray analysis identified 24 downregulated and 22 upregulated genes with a fold change of >2 (see tables 2 and 3), with a significant number of top-ranked genes being central to the IGF-1 and Wnt signalling pathways. IGF-1 was found to be significantly upregulated in TAO tissue, along with suppressor of cytokine signalling 3 (SOCS3) and serine/threonine-protein kinase-1 (SGK-1). Insulin-like growth factor binding protein 6 (IGFBP6) and insulin receptor substrate 2 were also found to be significantly downregulated.
Downregulated genes in orbital fat from active thyroid associated orbitopathy compared with normal control tissue, as analysed using microarrays
Upregulated genes in orbital fat from active thyroid associated orbitopathy compared with normal control tissue, as analysed using microarrays
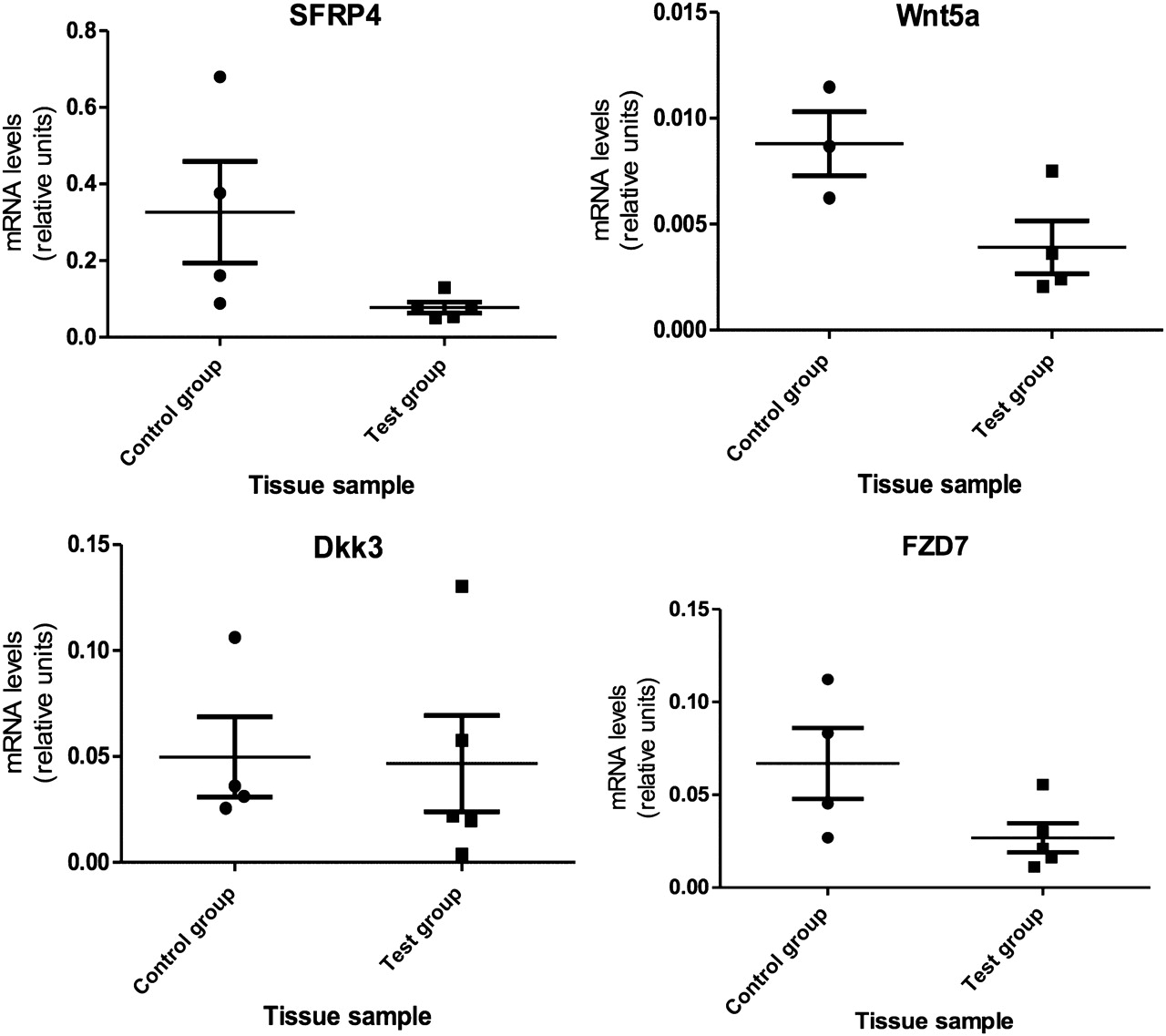
Most differentially expressed, Wnt signalling genes were downregulated, including Wnt5a, Dickkopf 3 (DKK3), sFRP4 and frizzled 7 (FZD7). Only dishevelled-associated activator of morphogenesis 1 (DAAM1) was found to be upregulated. Quantitative PCR was performed on nine genes, thus confirming the microarray findings (see figures 1 and 2).

Relative expression levels of IGF-1 signalling genes in control and test groups. IGF-1, p=0.039; 95% CI −0.035 to 0.0023; IRS2, p=0.006; 95% CI 0.018 to 0.11; SOCS3, p=0.014; 95% CI −0.16 to −0.012; SGK1, p=0.040; 95% CI −0.16 to 0.012; JUN, p=0.055; 95% CI 0.043 to 0.0057.
{kind=link}
{kind=link}
Relative expression levels of Wnt signalling genes in control and test groups. SFRP4, p=0.036; 95% CI −0.02950 to 0.5273; Wnt5a, p=0.027; 95% CI −0.00014 to 0.0099; DKK3, p=0.460; 95% CI −0.069 to 0.076; FZD7, p=0.037; 95% CI −0.0049 to 0.085.
Discussion
This investigation has demonstrated significant changes in the expression of Wnt signalling genes that may be relevant in the pathophysiology of TAO and confirms previous findings. Kumar et al13 examined 20 TAO and 8 control specimens of orbital fat and focused on sFRP-1, an inhibitor of Wnt signalling, which was found to be upregulated in active TAO tissue as a factor mediating adipogenesis; the hypothesis being confirmed by inducing adipocyte differentiation in orbital fibroblasts using sFRP-1. sFRP dysregulation has also been suggested in non-microarray single gene comparisons between active TAO orbital tissue and control tissue where no difference in SFRP-1 expression was identified, although SFRP-3 was found to be downregulated in active TAO tissue.14 The findings in the two other smaller gene array studies,15 ,16 identifying upregulation of adipocyte immediate early genes and CLN 2,3, were not confirmed by our study.
The findings of these previous microarray studies6 ,15 ,16 may not, however, reflect common patterns of gene expression in TAO, as the inclusion of a high proportion of patients with severe disease and optic neuropathy implies selection bias because tissue was sampled from patients at the most severe extreme (and minority) of the disease spectrum. The patients in this study with mild active disease represent the much more common phenotype of TAO. These differences in clinical selection may also explain the differences in the magnitude of fold change between this and previous microarray studies with higher fold changes observed in tissue derived from very severe cases. In addition, many of the tissues analysed were from patients receiving high-dose steroids, often for long periods of time, as well as radiotherapy; such treatments are likely to have modulated the immunopathological interactions underlying the disease and the associated gene expression pattern. By contrast, all the samples included in this study were from patients with mild active orbital disease that had not been treated. Furthermore, these previous microarray studies employed older cDNA microarray platforms covering a limited number of gene sets, whereas this study has used the gold standard Affymetrix Human Genome U133 Plus 2.0 array platform which offers a much wider genome-level transcription set covering 38 500 genes.
Wnt signalling may be involved in the pathogenesis of TAO
Wnt signalling genes are a complex family of growth regulators that govern a wide variety of tissue functions including maintenance, remodelling and differentiation processes.19 Wnts exert their effects through multiple so-called canonical and non-canonical pathways.
The canonical Wnt signalling cascade converges on the transcriptional regulator β-catenin. Wnt's binding to both frizzled (FZD) receptors and low density lipoprotein-related receptor co-receptors leads to the activation of cytoplasmic β-catenin; β-catenin is then translocated to the nucleus, where it binds to the Lymphoid enhancer-binding factor/transcription factor (LEF/TCF) family of transcriptional factors to activate Wnt effector genes (such as DAAM1). DAAM1 is an important regulator of actin dynamics, cellular adhesion and motility.20 The non-canonical pathway is poorly understood, but it is thought that Wnt and FZD interactions trigger intracellular calcium release, activating the phosphatase, calcineurin and the calcium-sensitive kinases CAMKII and protein kinase C; the downstream pathways of non-canonical signalling are yet to be elucidated.21 Wnt5a (downregulated in TAO; table 3) is classified as a protein activating the non-canonical pathway,22 although it has recently been demonstrated that Wnt5a can inhibit or stimulate the β-catenin pathway, depending on the receptor context.23
Wnt signalling has been widely shown to be important in the process of adipogenesis,21 although control of adipogenesis through Wnt signalling is complex. Canonical Wnt signalling is known to inhibit adipogenesis by blocking induction of PPARG and is thought to act as a molecular switch that represses adipogenesis.24 Consideration of the non-canonical pathway, however, adds complexity to this model as stimulation of the non-canonical pathway induces adipogenesis by antagonising the canonical Wnt/β-catenin pathway.21 In addition to Wnt ligands and receptors, secreted antagonists of Wnt–ligand binding have been identified,25 which include sFRPs, one of which (sFRP4) has been shown to be downregulated in this study (table 3).
As is evident from this description, Wnt signalling acts through multiple ligands, receptors, soluble inhibitors and across mutually antagonistic pathways creating an extremely complex system. This complex regulation of the Wnt signalling pathways makes the findings from this study difficult to interpret mechanistically. However, our findings identify, in line with previous studies,13 ,14 Wnt signalling genes as being differentially regulated in the orbital fat of untreated TAO, suggesting that further investigations into these signalling components may inform the pathogenesis of this condition.
IGF-1 signalling as a target for TAO treatment
The role of IGF-1 in TAO has been of much recent interest. IGF-1 exerts multiple actions on growth and metabolism,26 and IGF-1 signalling has also been implicated in carcinogenesis, where IGF-1 has been shown to inhibit apoptosis and encourage cellular proliferation.27 Recent investigations also strongly implicate IGF-1R as playing a central role in TAO and the specific activation of this receptor induces HA synthesis,9 whereas activation of the TSHR with TSH failed to mimic these responses. Co-localisation of TSHR and IGF-1R in orbital fibroblasts10 also lends support for the role of IGF-1 in the pathogenesis of TAO. In addition to its role in HA synthesis, IGF-1 is also strongly implicated in adipogenesis28 and may, therefore, regulate the transdifferentiation of orbital fibroblasts into adipocytes in this condition.
This investigation has clearly identified changes in the expression of several genes involved in IGF-1 signalling within orbital fat from active and untreated TAO patients, this being of particular relevance as fully humanised IGF-1R blocking drugs are a therapeutic reality and currently undergoing trials in oncology.29
This study has identified dysregulation of numerous genes involved in the IGF-1 signalling pathway. IGF-1 is the binding ligand that stimulates IGF-1R, which has also been shown to be upregulated in this study. IGF-1R is a transmembrane tyrosine kinase receptor and signalling transduction relies on essential IGF-1R binding/signalling genes, such as SOCS3 and insulin receptor substrate 2.30 SOCS3 is a negative regulator of IGF-1R signalling and it is thought that SOCS3 binds to IGF-1R and may be a direct substrate for the receptor tyrosine kinase.31 Downstream signalling and transcriptional regulators, such as SGK-1, are known to be important regulators of IGF-1—acting through PDK/Akt signalling32—and these factors were also shown to be upregulated in our investigation.
The exceptional rarity of untreated but active TAO tissue means that the low number of patient samples may be a limitation of this study. However, this study underscores the ability to obtain data even in rare diseases where tissue availability is exceptionally limited. Despite this limitation in sample size, this investigation has, for the first time, identified significant changes in two important molecular signalling systems in the orbital fat from patients with active and untreated TAO. First, we have confirmed previous reports of altered Wnt signalling,6 ,14 but further studies are warranted to identify how these genes regulate adipogenesis and HA synthesis in TAO, either through the canonical (β-catenin dependent) or non-canonical Wnt signalling pathways. Second, the demonstration of IGF-1 pathway dysregulation in TAO—being the first such in human tissues—is of major importance in the context of recent work identifying IGF-1R as a potential target for treating this condition5 and as humanised IGF-1R blocking drugs are currently being evaluated in clinical trials. In the absence of a functional animal model for TAO, these findings provide important evidence to support the use of IGF-1R antagonists in clinical trials for TAO.
Take-home messages
IGF-1 signalling genes are dysregulated in orbital fat tissue from patients with thyroid associated orbitopathy. This represents the first evidence from human tissue for IGF-1 dysregulation and provides further support for clinical trials of IGF-1 receptor antagonists for this condition.
References
Footnotes
Funding This study was funded bya Society for Endocrinology early career award (DGE) and Academy of Medical Sciences and Wellcome clinical lecturer starter grant (DGE). The authors acknowledge a proportion of their financial support from the Department of Health through the award made by the National Institute for Health Research to Moorfields Eye Hospital NHS Foundation Trust and UCL Institute of Ophthalmology for a Specialist Biomedical Research Centre for Ophthalmology. The views expressed in this publication are those of the authors and not necessarily those of the Department of Health.
Competing interests None.
Patient consent Obtained.
Ethics approval Moorfields Eye Hospital research governance committee.
Provenance and peer review Not commissioned; externally peer reviewed.