Article Text

Abstract
Aims Vulvar squamous cell carcinoma (VSCC) spreads early and mainly locally via direct expansion into adjacent structures, followed by lymphatic metastasis to the regional lymph nodes (LNs). In the lymphatic metastasis, cancer cells bearing CXCR4 and ACKR3 (CXCR7) receptors are recruited to the LNs that produce the CXCL12 ligand. Our study aimed to assess the role of the CXCR4/ACKR3/CXCL12 axis in VSCC progression.
Methods Tumour and LN tissue samples were obtained from 46 patients with VSCC and 51 patients with premalignant vulvar lesions. We assessed CXCR4, ACKR3 and CXCL12 by immunohistochemistry (IHC) in the tissue samples. Additionally, CXCL12 levels were determined by ELISA in the sera of 23 patients with premalignant lesions, 37 with VSCC and 16 healthy volunteers.
Results CXCR4 and ACKR3 proteins were virtually absent in vulvar precancers, while in VSCC samples the IHC staining was strong. In the LNs of patients with VSCC, 98% of metastatic cells expressed CXCR4 and 85% expressed ACKR3. Neither CXCR4 nor ACKR3 presence was correlated with tumour human papilloma virus status. Few CXCL12-positive cells were found in the analysed tissue samples, but serum CXCL12 levels were significantly increased in both patients with premalignant vulvar lesions and with VSCC compared with healthy volunteers.
Conclusions It appears that during progression and lymphatic spread of VSCC, the CXCR4/ACKR3/CXCL12 axis is activated. Moreover, our data suggest that CXCR4 antagonists merit further attention as a possible therapeutic option in patients with VSCC.
- biomarkers
- tumour
- blood proteins
- carcinoma
- cytokines
- female urogenital diseases
Data availability statement
Data are available upon reasonable request from the corresponding author.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Introduction
Vulvar squamous cell carcinoma (VSCC) is a rare disease accounting for about 4% of all gynecological malignancies. VSCC arises via human papilloma virus (HPV)-dependent and HPV-independent pathways, with approximately one-third of VSCC tumour cases being HPV-positive.1 2 In the first pathway, VSCC originates from the progression of high-grade squamous intraepithelial lesions (HSILs) associated with HPV infection.3 In the HPV-independent pathway, VSCC derives from differentiated-type vulvar intraepithelial neoplasia (dVIN).3 VSCC spreads early and mainly locally via direct expansion into adjacent structures, followed by lymphatic metastasis to the regional lymph nodes (LNs)—from the inguinal to the femoral and then to the pelvic LNs.4 5 Distant metastases from vulvar cancer are infrequent events.6
Lymphatics represent anatomical routes for tumour spread. Primary tumours promote lymphatic vessel formation (lymphangiogenesis), resulting from upregulation of a number of lymphangiogenic factors, for example, vascular endothelial growth factor C and vascular endothelial growth factor D, found to be correlated with the enhanced lymphatic tumour spread.7 Moreover, non-functional tumour lymphatics generate locally elevated interstitial fluid pressure, which is associated with an increased incidence of metastases.8 Besides the intratumoral lymphatic vessels, peritumoral lymphatics were also demonstrated to promote lymphogenic tumour spread.7 9 Abundant peritumoral lymphatic vessels were found to be associated with nodal metastases and shorter disease-free and overall survival times.10
Apart from their passive role in the metastatic spread, lymphatic vessels were also shown to contribute to the efficiency of the immune recognition and to the regulation of the checkpoints critical in antitumour responses. The LN lymphatic endothelial cells (LECs) may contribute to immune tolerance to T cells by programmed death-ligand 1 (PD-L1, CD274) expression.11 Thus, lymphatics play diverse, active roles in tumour progression.7 12 One of the mechanisms of the ‘active’ contribution of lymphatics to metastasis refers to a cross-talk between lymphatic and tumour cells. As shown in many human cancers and some experimental models, the lymphatic system promotes migration of cancer cells towards lymphatics by chemokine secretion.13 Cancer cells bearing specific receptors, such as C-X-C motif chemokine receptor 4 (CXCR4) and atypical chemokine receptor 3 (ACKR3, previously termed CXCR7), are recruited to the LNs that secrete a ligand of these receptors, C-X-C motif chemokine ligand 12 (CXCL12 chemokine, previously termed SDF-1), produced by the LECs. This means that, in order to disseminate, cancer cells use the physiological signalling mechanism of attracting antigen-presenting cells to the LNs.9 14 LNs, similar to lungs, bone marrow and liver, all known to constitutively express CXCL12, are the common sites of metastases.15 In addition, to migrate toward the draining lymphatics, tumour cells, along with expressing ACKR3, may often also secrete the ACKR3 ligands, and—by exploiting interstitial flow—produce an autologous chemotaxis-inducing gradients.13 16 Interestingly, the CXCR4/ACKR3/CXCL12 axis was previously shown to activate the PI3K/Akt/mTOR pathway,15 17 which we demonstrated to be activated in VSCC.18 19
CXCL12 belongs to the chemokines inducing the neogenesis of tertiary lymphoid structures (TLSs), the LN-like formations present in the microenvironment of chronic inflammation sites in many solid tumour types.20 TLSs are associated with favourable patient prognosis and may be exploited in several strategies of reversing immune tolerance and increasing the antitumour immune response, that is, using immune checkpoint inhibitors (ICIs).20 21 Although there are no data on TLS in VSCC, high infiltration by T cells22 and the expression of PD-L123 in vulvar tumours suggest that VSCC might be amenable to ICI. In VSCC, PD-L1 positivity of peritumoural immune cells and of cancer cells are independent prognostic factors of favourable and poor outcome, respectively.24 25 Solid tumours that contain high numbers of tumour-infiltrating lymphocytes are classified as hot tumours and are most likely to be highly responsive to ICI. Yet, very preliminary data on ICIs in advanced or recurrent VSCC reveal modest efficacy.23 Apart from measuring PD-L1 expression of tumour cells and lymphocyte infiltration, the outcome of immune checkpoint inhibition can be predicted by tumour mutational burden (TMB), and high TMB is another feature of hot tumours. However, based on a recent molecular profiling26 VSCC should be classified as TMB-low type of cancer. CXCL12 immunosuppressive activity15 is among other possible reasons for so far limited success of anti-PD-L1 therapy.
Activation of CXCR4/ACKR3/CXCL12 axis causes numerous biological pleiotropic effects. CXCL12 and its cognate receptor CXCR4 do not only induce metastatic outgrowth in the LNs but also contribute to the premetastatic niche formation by recruiting immunosuppressive cells to the premetastatic LN.27 The lymphatic system is the main pathway for metastatic spread of VSCC, and thus our study aimed to assess the role of CXCR4 and ACKR3 and their main ligand, CXCL12, in the progression of VSCC.
Materials and methods
Patients
Clinical material was obtained from patients undergoing surgery because of VSCC, patients with premalignant vulvar lesions, that is, HSIL and dVIN (tissue samples and sera) and from healthy volunteers (sera) at the Maria Sklodowska-Curie National Research Institute of Oncology in Warsaw and at the Holycross Cancer Center in Kielce, Poland, between January 2002 and January 2018. The numbers of patients, according to the methods applied in the study for the respective samples, are provided in table 1.
Basic demographic data and numbers of patients and healthy volunteers enrolled
HPV genotyping
The HPV status was determined, as described previously,18 using the AmpliSens HPV HCR-genotype-titre-FRT kit (InterLabService), which detects 14 high-risk human papilloma virus (hrHPV) genotypes, namely, HPV16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 66 and 68, following the manufacturer’s instructions.
Immunohistochemistry (IHC)
Immunohistochemical staining was performed on 4 µm formalin-fixed paraffin-embedded (FFPE) tissue sections using the EnVision FLEX+, Mouse, High pH Detection System (Dako, Glostrup, Denmark). Sections were deparaffinised with xylene and rehydrated in ethanol solutions. Heat-induced epitope retrieval was carried out in Target Retrieval Solution (Dako) for 25 min at 96°C. After cooling, the slides were treated for 5 min with an endogenous peroxidase blocker (Dako). Slides were incubated overnight at 4°C with monoclonal antibodies against CXCR4 (dilution 1:1000, UMB2; Abcam), ACKR3 (dilution 1:500, anti-GPCR RDC1, Abcam) and CXCL12/SDF-1 (dilution 1:50, D8G6H; Cell Signalling). The colour reaction products were developed with the Envision Detection System, Peroxidase/DAB+, Rabbit/Mouse (Dako, Denmark). Nuclear contrast was achieved with haematoxylin counterstaining. Positive tissue controls for anti-CXCR4, ACKR3 and CXCL12 staining consisted of normal colon mucosa and additionally for CXCL12—of breast cancer tissue.
Analysis of CXCR4 and ACKR3 immunostaining outcomes in vulvar tissues was performed using optical microscopy (×400) and the results were assessed on a four-grade scale of staining intensity (0—none, 1—weak, 2—moderate and 3—strong for staining). Histoscore (H-score) was used as a method of assessing the extent of immunoreactivity and calculating the percentage of cells at each staining intensity level according to the following formula:
H-score=[1×(% cells 1)+2×(% cells 2)+3×(% cells 3)]. The range of possible scores is from 0 to 300.
Diagnosis of premalignant lesions (dVIN and HSIL) was based on routine H&E staining, along with p16 and TP53 IHC, as described previously18 and in compliance with current WHO 2020 recommendations.28
ELISA quantification of serum CXCL12
Serum samples were obtained from blood of 19 patients with premalignant vulvar lesions (6 with dVIN and 13 with HSIL), 45 patients with VSCC and 16 healthy volunteers (median age 45, range 25–64 years). All sera were separated within 1 hour after blood collection and stored at −70°C until assayed, equally for the study and control groups. Serum CXCL12 levels were determined using human SDF-1 (stromal cell derived factor 1) ELISA Kit (Wuhan Fine Biotech Co.), according to the manufacturer’s instructions.
Statistical analysis
Kruskal-Wallis test followed by Mann-Whitney multiple pairwise comparisons was used to test the significance of differences in protein expression between the tissue and serum sample groups. Benjamini-Hochberg procedure was used to adjust p values due to multiple hypothesis testing. The association between hrHPV status of the tumours and CXCR4 and ACKR3 protein expression was assessed using Mann-Whitney test. Spearman correlation coefficient and AS89 algorithm were used to inspect correlation of selected variables. The association of CXCR4 and ACKR3 protein expression with survival of patients with primary VSCC was determined using Cox proportional hazard model implemented in the survival package (V.3.1–8) in the R environment V.3.6.3 (http://www.R-project.org). The p value of <0.05 was considered significant. The results were visualised using GraphPad Prism (La Jolla, California, USA).
Results
CXCR4 and ACKR3 staining in VSCC and its premalignant lesions
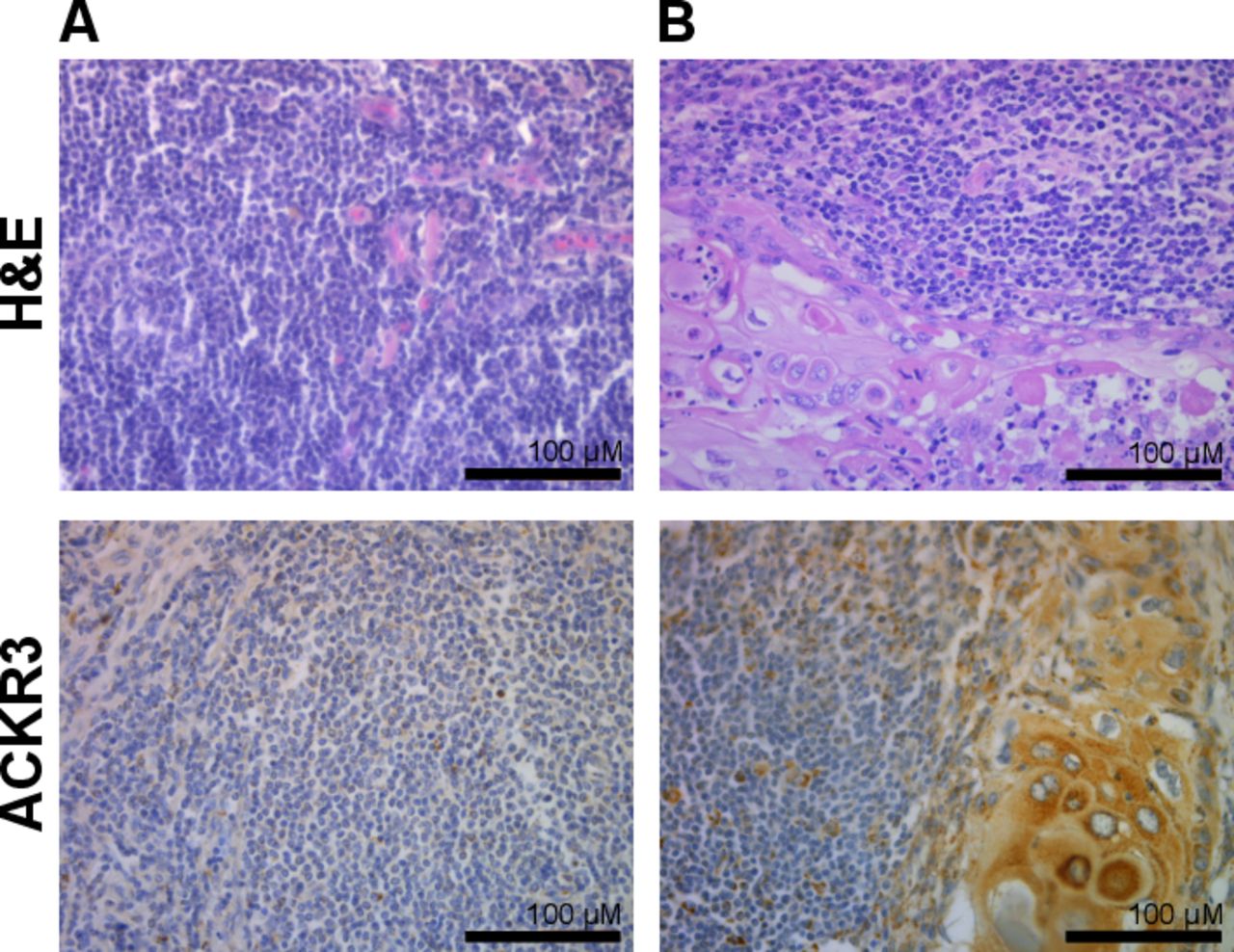
The expression of CXCR4 and ACKR3 receptors was determined by IHC analyses of these proteins on FFPE tissue specimens obtained from patients with VSCC and those with premalignant lesions. The samples of premalignant lesions included HSIL, which is related to hrHPV (predominantly type 16) infection and dVIN, the hrHPV-independent precursor of VSCC. The diagnosis of these lesions of the vulva based on histological criteria was confirmed using anti-p16 and p53 IHC. CXCR4 staining was negative or weak in premalignant lesions, whereas in VSCC tissues it was strong and both membranous and cytoplasmic (figure 1, middle panel). ACKR3 staining was weak in premalignant lesions, but in VSCC tissues it was strong, cytoplasmic, with clearly visible granularities (figure 1, right panel). In general, significantly higher numbers of lesion and cancer cells were CXCR4-positve than ACKR3-positive. Both proteins, CXCR4 and ACKR3, were significantly more abundant in tumour cells (both lesion and cancer cells) compared with infiltrating lymphocytes. In virtually all of the examined cases, CXCR4 and ACKR3 staining was negative in histologically normal vulvar tissue adjacent to the tumour.

Examples of H&E (left panel), immunohistochemical CXCR4 (middle panel) and immunohistochemical ACKR3 (right panel) staining performed on tissue sections of premalignant (A,B) and VSCC (C–F) tumours. (A) HSIL sample, (B) dVIN, (C,D) VSCC samples obtained from patients with no progression, (E) VSCC sample obtained from a patient who experienced subsequent disease progression, (F) VSCC recurrence, (G) positive control (colon mucosa sample). Left panel: H&E-stained specimen, middle panel: CXCR4 immunostained sections, right panel: ACKR3 immunostained sections. Images were taken at ×40 magnification. Scale bar, 100 µm. ACKR3, atypical chemokine receptor 3; CXCR4, C-X-C motif chemokine receptor 4; dVIN, differentiated-type vulvar intraepithelial neoplasia; HSIL, high-grade squamous intraepithelial lesion; VSCC, vulvar squamous cell carcinoma.
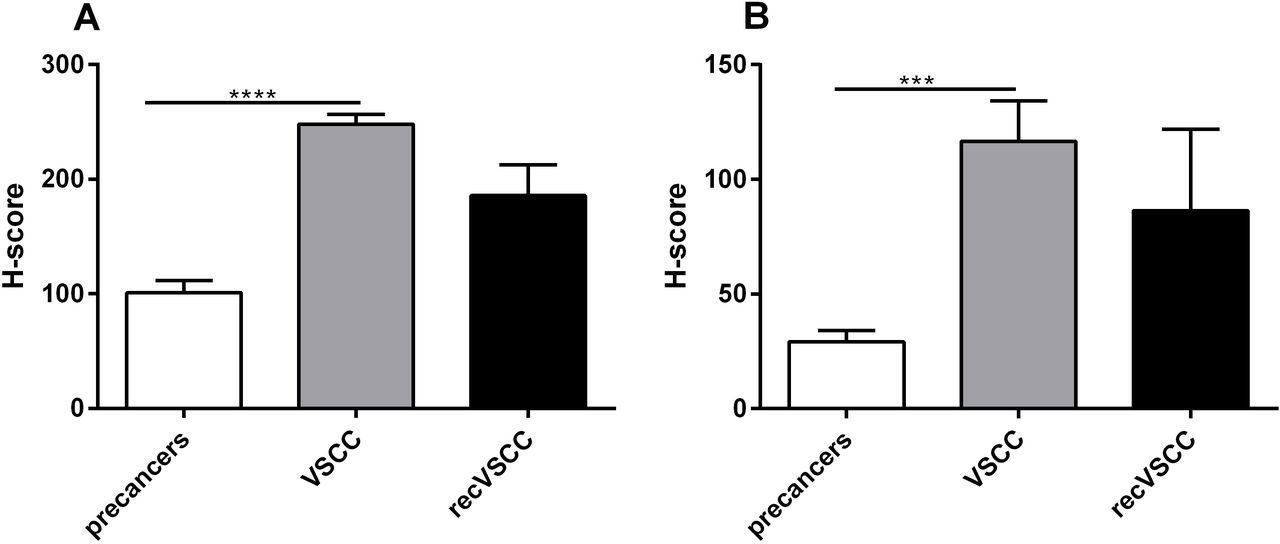
IHC results were evaluated using a semiquantitative approach by assigning an H-score to protein expression patterns within the lesion and cancer cells as well as infiltrating lymphocytes in vulvar premalignant lesions (HSIL and dVIN), primary VSSC and recurrent VSCC tumours. The results of such IHC analysis, taking into consideration both the staining intensity and the percentage of the stained cells, are depicted in figures 2 and 3 for CXCR4 and ACKR3, respectively. The abundance of both proteins proved to be significantly higher in VSCC compared with vulvar premalignant lesion samples, suggesting the increasing expression of CXCR4 and ACKR3 with VSCC development.

CXCR4 H-score distribution for neoplastic cells (A) and lymphocytes (B) in premalignant lesions (HSIL n=36 and dVIN n=13), primary VSCC (n=37) and recVSCC (n=8) tumours. ***p value ≤ 0.001; ****p value ≤ 0.0001. CXCR4, C-X-C motif chemokine receptor 4; dVIN, differentiated-type vulvar intraepithelial neoplasia; H-score, histoscore; HSIL, high-grade squamous intraepithelial lesion; recVSCC, recurrent vulvar squamous cell carcinoma; VSCC, vulvar squamous cell carcinoma.
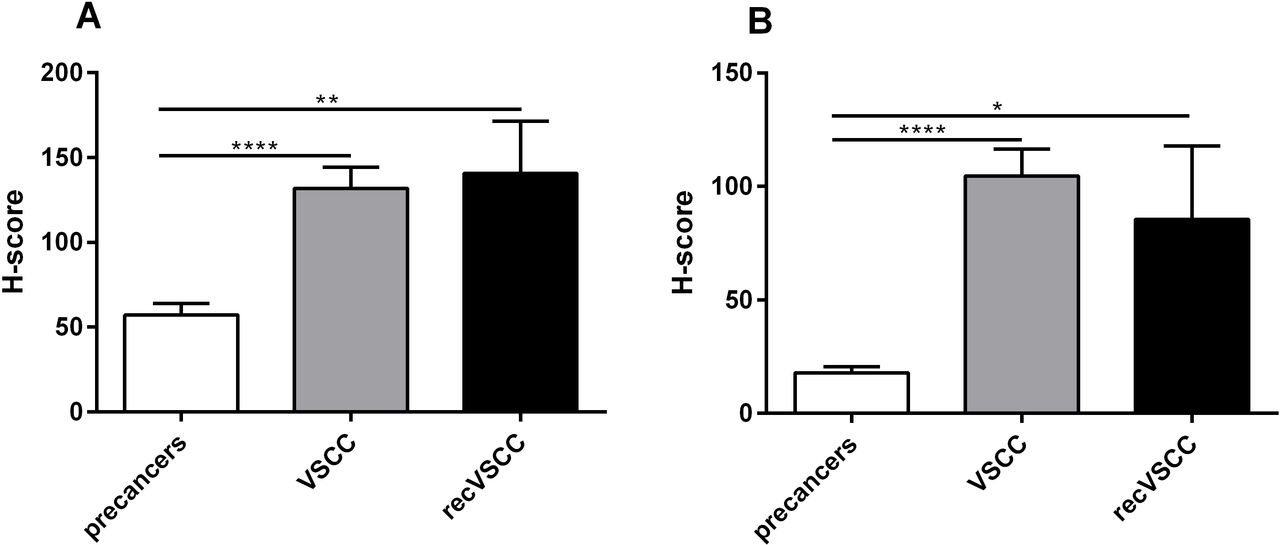
ACKR3 H-score distribution for neoplastic cells (A) and lymphocytes (B) in premalignant lesions (HSIL n=36 and dVIN n=13), primary VSCC (n=38) and recVSCC (n=9) tumours. *p value ≤ 0.05; **p value ≤ 0.01; ***p value ≤ 0.001; ****p value ≤ 0.0001. ACKR3, atypical chemokine receptor 3; dVIN, differentiated-type vulvar intraepithelial neoplasia; H-score, histoscore; HSIL, high-grade squamous intraepithelial lesion; recVSCC, recurrent vulvar squamous cell carcinoma; VSCC, vulvar squamous cell carcinoma
The ratios between the ACKR3 H-score results for lesion or cancer cells versus the H-score results for lymphocytes revealed the decreasing contribution of the ACKR3-stained tumour cells compared with lymphocytes during VSCC progression (figure 4B). In contrary to ACKR3, the relative CXCR4 abundance in cancer cells seems to be increasing compared with the lymphocytes during VSCC, although this observation is less apparent (figure 4A). There was no significant correlation between ratios of cancer to lymphocytes between CXCR4 and ACKR3 (r=0.29, p=0.08). Spearman correlation analysis revealed a significant association between cancer and lymphocyte expression for both receptors, CXCR4 (r=0.43, p=0.007) and ACKR3 (r=0.52, p=0.0009). Additionally, there was a positive correlation between the expression of CXCR4 and ACKR3 in cancer cells (r=0.34, p=0.045).

CXCR4 (A) and ACKR3 (B) H-score ratios for neoplastic cells versus lymphocytes in premalignant lesions (HSIL n=36 and dVIN n=13), primary VSCC (n=38) and recVSCC (n=8) tumours. ***p value ≤ 0.001. ACKR3, atypical chemokine receptor 3; CXCR4, C-X-C motif chemokine receptor 4; dVIN, differentiated-type vulvar intraepithelial neoplasia; H-score, histoscore; HSIL, high-grade squamous intraepithelial lesion; recVSCC, recurrent vulvar squamous cell carcinoma; VSCC, vulvar squamous cell carcinoma.
As HPV was previously shown to affect CXCR4 levels,29 we investigated correlations between the presence of hrHPV and the percentages of CXCR4 and ACKR3-positive cancer cells in the examined VSCC tumours. Unexpectedly, the Mann-Whitney test revealed no relationship between the hrHPV status of VSCC tumours and CXCR4 or ACKR3 protein expression, neither in cancer cells nor in lymphocytes (data not shown).
CXCR4 and ACKR3 staining in LNs of patients with VSCC
Once we proved that CXCR4 and ACKR3 proteins were abundant in VSCC tumours, we checked whether these receptors contribute to cancer cell recruitment to LNs. Examples of immunohistochemical CXCR4 and ACKR3 staining performed on LNs obtained from patients with VSCC are depicted in figures 5 and 6, respectively. The receptor-positive metastatic cells were abundant in the involved LNs, with 98% of positive cells for CXCR4 and 85% for ACKR3 (H-score results depicted in online supplemental figure 1A). CXCR4-positive and ACKR3-positive lymphocytes were also found in both involved (50% and 45%, respectively) and metastasis-free LNs (50% and 15%, respectively) (H-score results presented in online supplemental figure 1B).
Supplemental material

Examples of immunohistochemical CXCR4 staining performed on LNs obtained from patients with VSCC. Sections of metastasis-free LN (A) and involved (B) LN. Upper panel : H&E-stained specimen, lower panel: CXCR4 immunostained sections. Images were taken at ×40 magnification. Scale bar, 100 µm. CXCR4, C-X-C motif chemokine receptor 4; LN, lymph node; VSCC, vulvar squamous cell carcinoma.

Examples of immunohistochemical ACKR3 staining performed on LNs obtained from patients with VSCC. Sections of metastasis-free LN (A) and the involved (B) LN. Upper panel: H&E-stained specimen, lower panel: CXCR4 immunostained sections. Images were taken at ×40 magnification. Scale bar, 100 µm. ACKR3, atypical chemokine receptor 3; CXCR4, C-X-C motif chemokine receptor 4; LN, lymph node; VSCC, vulvar squamous cell carcinoma.
CXCL12 staining in vulvar tumours and its LNs of patients with VSCC
CXCL12 protein expressed in the LNs recruits cancer cells bearing CXCR4, the CXCL12 receptor, directly contributing to metastasis formation. We performed IHC staining on VSCC tumour and LN tissue sections with an antibody against CXCL12. Only scant numbers of CXCL12-positive cancer and lymphocytes within preinvasive vulvar lesions, VSCC tumours and LNs of patients with VSCC were identified (figure 7 and online supplemental figure 2). However, CXCL12 protein was present in blood vessels of nearly all of the preinvasive vulvar lesions and VSCC tumours analysed.
Supplemental material

Example of immunohistochemical CXCL12 staining of blood vessels in VSCC tumour (A). Immunohistochemistry in positive control tissues: breast cancer (B) and colon mucosa samples (C). Upper panel: H&E-stained specimen, lower panel: CXCR4 immunostained sections. (A, lower panel) Positive immunohistochemical CXCL12 staining of blood vessels (thick arrows) and negative staining in carcinoma cells (thin arrows). Images were taken at ×40 magnification. Scale bar, 100 µm. CXCL12, C-X-C motif chemokine ligand 12; CXCR4, C-X-C motif chemokine receptor 4; VSCC, vulvar squamous cell carcinoma.
Prognostic role of CXCR4 and ACKR3 protein abundance for relapse-free survival of patients with VSCC
Next, the CXCR4 and ACKR3 expression was assessed for associations with FIGO (The International Federation of Gynecology and Obstetrics) stage and tumour grade (G) as well as with time to recurrence in patients with primary VSCC (n=37), using Cox proportional hazard model. The percentages of CXCR4 or ACKR3-positive cancer cells and lymphocytes did not correlate with FIGO stage or with grade. The relapse-free survival was found not to be correlated with any of the tested variables, that is, with the percentages of CXCR4 or ACKR3-positive cancer cells (p=0.21 and 0.95, respectively) or lymphocytes (p=0.67 and 0.24, respectively) in primary VSCC tumours. No correlation between the relapse-free survival and either FIGO (p=0.079) or G (p=0.157) was found; probably the number of samples was too low to assess the statistical significance in the context of patients’ survival.
Serum CXCL12 protein in patients with VSCC
CXCL12 concentrations in the sera obtained from patients with vulvar premalignant lesions (HSIL and dVIN) and patients with VSCC, as well as from healthy volunteers are shown in figure 8. Serum CXCL12 levels were found to be significantly higher in patients with vulvar precancers and in patients with VSCC than in controls. CXCL12 levels did not correlate with FIGO (p=0.14, r=−0.28) nor with grade (p=0.47, r=0.14). In patients with primary VSCC tumours (n=30), the relapse-free survival was not associated with serum CXCL12 levels (p=0.18 in the Cox proportional hazard model).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
CXCL12 levels assessed by ELISA in the sera of healthy volunteers (C, control samples, n=16) and patients with premalignant lesions (HSIL n=16 and dVIN n=7), VSCC (n=36) and recVSCC tumours (n=2). *p value ≤ 0.05. CXCL12, C-X-C motif chemokine ligand 12; dVIN, differentiated-type vulvar intraepithelial neoplasia; HSIL, high-grade squamous intraepithelial lesion; recVSCC, recurrent vulvar squamous cell carcinoma; VSCC, vulvar squamous cell carcinoma.
Discussion
Survival in VSCC is highly dependent on the size of the lesion and on the pathological status of the inguinal LNs, as this cancer spreads predominantly through the lymphatic vessels. Patients with histologically negative inguinal LNs have survival rates of 90% vs 50% for those with histologically positive inguinal LNs.4 Even in recurrent disease, survival rates are lower for patients with inguinal/pelvic than for those with vulvar recurrence, that is, 27% vs 60% at 5 years.5
The CXCR4/ACKR3/CXCL12 axis is a crucial mechanism regulating trafficking of cancer cells to the lymphatic sites of metastasis. CXCR4 and ACKR3 receptors ‘guide’ cancer cells to LNs that provide ligands for these receptors. CXCL12 is a common ligand for both CXCR4 and ACKR3; the ACKR3 receptor can also bind to CXCL11, although with a lower affinity.30 Because LN status is the most important determinant of survival in VSCC and the CXCR4/ACKR3/CXCL12 axis was shown to regulate mTOR signalling,17 which is upregulated in VSCC,18 19 we investigated the expression CXCR4 and ACKR3 proteins in VSCC tumours and in premalignant vulvar lesions. Both VSCC tumours and vulvar premalignant lesions expressed these receptors. CXCR4 staining was membranous and cytoplasmic, while ACKR3 showed cytoplasmic staining only. A previous in vitro study31 has shown that the internalisation of ACKR3, which is distributed in punctate intracellular vesicles, depends on the presence of CXCL12. The punctate cytoplasmic distribution of ACKR3 found in cultured cell lines incubated with CXCL1231 seems to correspond to the observed cytoplasmic ACKR3-stained granularities in VSCC tissues in our study.
We observed higher numbers of tumour cells being CXCR4-positve than ACKR3-positive. Both receptors, CXCR4 and ACKR3, were significantly more abundant in both lesion (HSIL and dVIN) and cancer cells than lymphocytes. To assess CXCR4 and ACKR3 expressions in cancer and stromal cells in more detail, we calculated the ratios between the H-scores for the two compartments. The results of this calculation suggest increasing contribution of CXCR4-positive cancer cells and decreasing of ACKR3-positive cancer cells compared with the receptor-stained lymphocytes during VSCC progression. Interestingly, there was a positive correlation between CXCR and ACKR3 expression in cancer cells, and the expression of both receptors positively correlated between cancer and lymphocytes, which confirms the complexity of CXCR and ACKR3 expressions’ regulation. A positive correlation between CXCR and ACKR3 expression may suggest CRCR7/ACKR3 hetorodimer formation32 in cancer cells strengthening the CRCL12 signal, yet this assumption requires verification in further research.
The CXCR4 and ACKR3 protein abundance was not found to correlate with hrHPV infection in VSCC tumours in our sample set. This was an unexpected finding, as some previously reported data showed an involvement of CXCL12 pathway in HPV-related cellular changes, that is, HPV was demonstrated to weaken cellular response to the CXCL12 stimulus,33 while E6 and/or E7 HPV proteins were shown to upregulate CXCR4 levels in HPV-associated cells by inducing its stabilisation.29
The presence of lymphovascular space invasion (LVSI) was also shown to be a strong predictor of LN metastasis and of recurrence and death in patients with VSCC.34–36 Tumour LVSI should be assessed during the routine histopathological workup not only for its prognostic value, but also—in patients with negative surgical margin—it might influence the choice of postoperative management.37 38 Lymphatic vessel density (LVD) has emerged as a promising prognostic indicator associated with LN involvement in several cancer types, including squamous cell carcinomas, with high intratumoural and peritumoral LVD being associated with poor and favourable prognosis, respectively.39–41 In patients with VSCC, an increased density of both blood and lymphatic microvessels was shown to be related to poor survival.42 However, in a more recent study,43 intratumoural LVD was unexpectedly found to be associated with a higher rate of overall survival and a lower rate of LN metastasis in patients with VSCC. As CXCR4/ACKR3/CXCL12 axis is known to orchestrate site-specific metastasis, we also investigated CXCL12 and its receptors’ expression in the regional LNs of patients with VSCC.
Although our study contains a small number of the regional LN cases of patients with VSCC, the high quantity of receptor-positive metastatic cells, for CXCR4 reaching nearly 100% and for ACKR3—85%—clearly suggests the role of CXCR4/ACKR3/CXCL12 axis in the lymphatic spread. The CXCR4-positive and ACKR3-positive lymphocytes were also found in the LNs, although at lower frequencies. As our data suggested that CXCR4 and ACKR3 receptors contributed to VSCC development and metastasis formation, we expected their common ligand, CXCL12, to be expressed in VSCC samples. However, we did not observe a significantly increased number of CXCL12-positive cells neither in preinvasive vulvar lesions and VSCC tumours nor in the LNs of patients with VSCC. The virtual absence of CXCL12 protein in VSCC tumours was unexpected as this chemokine was not only found in other tumour types, such as breast, ovarian or lung cancers, but was also shown to be of prognostic value.44–46 Our finding could be linked to a fast turnover of CXCL12 protein within the cells, due to an increased abundance of ACKR3, which mediates ligand internalisation and its subsequent degradation.47 48
By IHC, CXCL12 protein was detected in the blood vessels of nearly all of the analysed preinvasive vulvar lesions and VSCC tumours. CXCL12 on endothelial cells contributes to the induction of angiogenesis.15 49 The observed anti-CXCL12 staining of endothelia and vascular smooth muscle, similar to the staining pattern of anti-CD31 or CD34 antibodies, is consistent with the previous report of Pablos et al 50 showing both CXCL12 mRNA and protein to be expressed by normal endothelial cells. It is established that endothelial cells can secrete CXCL12,51 and thus it can by hypothesised that VSCC tumours contain soluble CXCL12 protein (undetectable by IHC). The source of high levels of circulating CXCL12 protein remains even more speculative. In the sera obtained from patients with vulvar premalignant lesions and those with VSCC, compared with those obtained from healthy volunteers, we showed significantly increased CXCL12 concentrations. Changes in CXCL12 receptor expression are known to contribute to the onset or progression of human diseases including cancer.52 As proinflammatory activities are attributed to CXCL12,52 its increased levels in patients with cancer sera may also reflect systemic inflammatory response that accompanies cancer progression.
For several cancer types, including cervical cancer,53 the expressions of CXCR4 and ACKR3 proteins have previously been shown to be associated with poor survival. Previously, Shiozaki et al 54 in their analysis of 30 primary invasive vulvar cancers, including 22 cases of VSCC, also reported poorer prognosis in patients with tumours that expressed CXCR4 compared with those with CXCR4-negative tumours. Probably, still too small patient group with VSCC in our study did not allow us to draw conclusions regarding the association of tumour expression of these receptors on disease-free survival. Larger series are needed to investigate the prognostic value of CXCR4 and ACKR3 protein expression.
In conclusion, the present study shows that he CXCR4/ACKR3/CXCL12 axis is activated during VSCC progression. Our results reveal that especially an increased expression of CXCR4 may accompany VSCC development and, being associated with the lymphatic spread of VSCC, may contribute to a more aggressive tumour phenotype. These findings suggest that the CXCR4/ACKR3/CXCL12 axis is vital for the metastatic spread in VSCC and supports the notion that CXCR4 antagonists55 merit further attention as a possible therapeutic option in patients with VSCC.
Take home messages
C-X-C motif chemokine receptor 4 (CXCR4)/atypical chemokine receptor 3 (ACKR3)/C-X-C motif chemokine ligand 12 (CXCL12) is the axis activated during the development and progression of vulvar squamous cell carcinoma (VSCC).
CXCR4 and ACKR3 proteins are virtually absent in vulvar precancers but abundant in VSCC tumours.
Metastatic cells express both CXCR4 and ACKR3 in the lymph nodes of patients with VSCC.
CXCL12 levels are significantly increased in patients with vulvar precancers and VSCC compared to healthy volunteers.
Data availability statement
Data are available upon reasonable request from the corresponding author.
Ethics statements
Patient consent for publication
Ethics approval
The study received ethical institutional board approvals of the Maria Sklodowska-Curie National Research Institute of Oncology (number 44/2002, 16/2015) and Holycross Cancer Centre (number 15/2014). All patients gave their informed consent to be included in the study, and all methods were performed in accordance with the relevant guidelines and regulations.
Acknowledgments
The authors gratefully acknowledge the assistance of Transition Technologies S.A. in patient data analysis using the MedStream Designer (MSD) application on the Onko.Sys platform.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Handling editor Mona El-Bahrawy.
NR and KK contributed equally.
Contributors MK contributed to the study conceptualisation and funding acquisition. NR, KK, SZ, BK, MF, JK and EB-Z contributed to experimental design and investigation. KZ and MB contributed to the collection and assembly of data from the patient materials. KG, AK and MK analysed the data. All authors were involved in writing the paper and gave the final approval of the submitted version.
Funding This work was financed by the Polish National Science Centre (grant number UMO-2013/10/E/NZ5/00663) and further supported by the Maria Sklodowska-Curie National Research Institute of Oncology subsidy from the Ministry of Science and Higher Education (including grant number GW36MB/2017).
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.