Article Text

Abstract
Basement membrane (BM) is an amorphous, sheet-like structure separating the epithelium from the stroma. BM is characterised by a complex structure comprising collagenous and non-collagenous proteoglycans and glycoproteins. In the breast, the thickness, density and composition of the BM around the ductal lobular system vary during differing development stages. In pathological conditions, the BM provides a physical barrier that separates proliferating intraductal epithelial cells from the surrounding stroma, and its absence or breach in malignant lesions is a hallmark of invasion and metastases. Currently, diagnostic services often use special stains and immunohistochemistry (IHC) to identify the BM in order to distinguish in situ from invasive lesions. However, distinguishing BM on stained sections, and differentiating the native BM from the reactive capsule or BM-like material surrounding some invasive malignant breast tumours is challenging. Although diagnostic use of the BM is being replaced by myoepithelial cell IHC markers, BM is considered by many to be a useful marker to distinguish in situ from invasive lesions in ambiguous cases. In this review, the structure, function and biological and clinical significance of the BM are discussed in relation to the various breast lesions with emphasis on how to distinguish the native BM from alternative pathological tissue mimicking its histology.
- BREAST
- Breast Neoplasms
- CARCINOMA
- COLLAGEN
Statistics from Altmetric.com
Introduction
In the normal breast tissue, the duct lobular system comprises two cell types, epithelial and myoepithelial cells (MECs), which are separated from the surrounding stromal tissue by a thin, sheet-like structure layer called the basement membrane (BM) (figure 1). MECs, which are located between the BM and the epithelial cells, play key roles in synthesising BM components in normal tissue. MECs secrete laminin 1, which is the major component of the BM, and synthesise other BM components including collagen IV, laminin 5, nidogen and fibronectin. MECs produce matrix metalloproteinases, which facilitate BM remodelling.1 MECs also possess BM receptors, including integrins, which mediate cell-BM attachment. MECs attach to BMs via hemidesmosomes and to the adjacent myoepithelial and luminal epithelial cells using desmosomes. MECs in normal ducts form a continuous layer while those in the terminal duct lobular units (TDLUs) are discontinuous, allowing some luminal epithelial cells direct contact with the BM.2
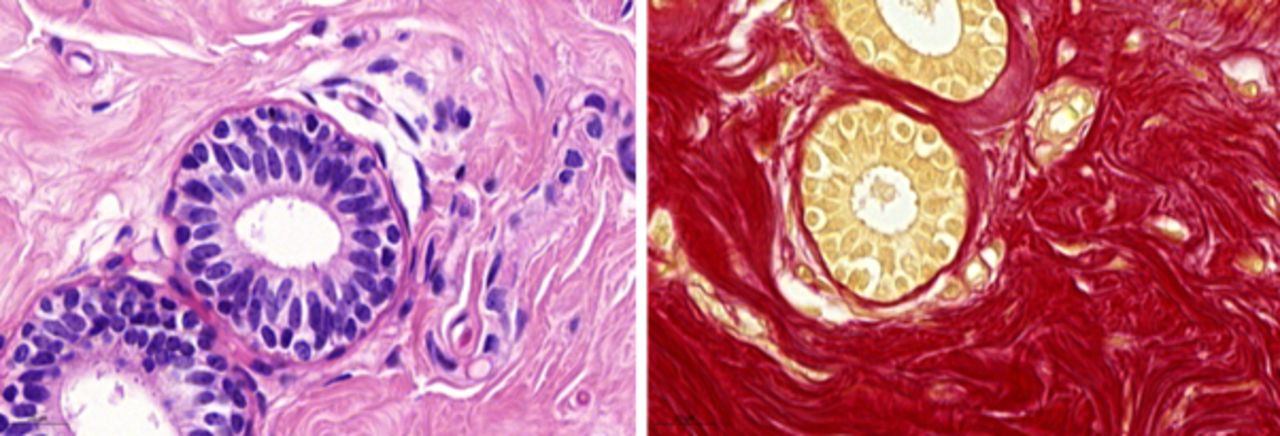
Photomicrographs of normal breast tissue basement membranes (BM). (A) H&E showing a well-defined BM that surrounds the glands. (B) Picaro Sirius Red (PSR) staining shows the BM as a red definite layer surrounding a normal duct (magnification ×63). Scale bar=20 µm.
BMs are thin, pliable and amorphous sheet-like structures with an extracellular matrix (ECM) component that provides cell support and acts as a platform for complex signalling. BMs comprised collagenous and non-collagenous components such as proteoglycans (PG) and glycoproteins.3 However, BMs undergo dynamic transformations throughout life, and the molecular structure and arrangement of BM fibres differ in various tissues according to their functions and between normal, benign and malignant lesions.4 In malignant lesions, the normal production and assembly of the BM is disrupted. In fact, BM changes further define the tumour microenvironment and provide host-derived regulatory signals during tumour progression. Animal models have indicated that destruction of the BM is associated with genetic instability and tumourigenesis.5
In the breast, the thickness, density and composition of the BM around the normal duct lobular system (termed native BM) may vary slightly during different development stages.6 In pathological conditions, including benign and malignant in situ lesions, the BM is typically preserved, but often exhibits altered thickness, structure or continuity. In malignant breast lesions, the preservation of the BM is used to indicate the in situ nature of the tumours (eg, ductal carcinoma in situ (DCIS) or lobular carcinoma in situ) whereas the absence or breach of the BM denotes invasion.7
In diagnostic practice, breast pathologists tend to use special stains and/or immunohistochemistry (IHC), rather than H&E staining, to distinguish one or more of the BM components that differentiate between in situ and invasive tumours in doubtful cases. Despite the advances in BM detection, distinguishing between native BM and reactive BM-like structures or pseudocapsule surrounding some malignant breast tumours remains problematic.7 Indeed, some invasive tumours show BM-like material, and it is known that the BM structure shares some components with the ECM of the interstitial stromal tissue and scar tissue, further complicating the differentiation between these types of tissues.8 In addition, the expression of some of the BM components by invasive breast tumours makes interpretation of the findings challenging.9 10
Although the diagnostic use of BM is now being replaced by MEC IHC markers in routine breast pathology practice, some pathologists consider the demonstration of a BM a specific finding, useful for distinguishing in situ from invasive lesions.7 The presence of a BM-like reactive capsule around encapsulated papillary carcinoma (EPC) is also used as the main evidence to support its in situ nature.11 In addition, the demonstration of BM in microglandular adenosis (MGA) is used to indicate its benign nature despite the lack of MECs, proliferating epithelial cell clonality and its infiltrative appearances.12 Therefore, the diagnostic use of BMs to classify breast lesions remains controversial. In this review, we discuss the structure, function and biological and clinical significance of BMs in various breast lesions.
Composition of the BM
By electron microscopy (EM), the BM is well organised and comprises three layers: (1) a linear, homogeneous, electron-dense region (lamina densa) and (2) a clear zone (lamina lucida) directly beneath MECs, and epithelial cells within the breast ducts and lobules. These two layers constitute the basal lamina, which is located towards the MECs. (3) The outer layer of BM is called reticular lamina, or lamina fibroreticularis, and is formed mainly of collagen III.13–15
The lamina densa is known as the BM proper and is formed from densely packed fibrils embedded in dense matrix.16 The lamina densa is between 20 and 300 nm thick, consisting of a network of collagen IV fibrils with PG perlecan, while the lamina lucida is ~40 nm thick and consists of laminin, nidogens (entactin) and integrins. A bridge of anchoring filaments of collagen VII connects the lamina densa and lucida. Hemidesmosomes, which are present on the basal aspect of MECs, are connected to the adjacent basal lamina by fine filaments.13–15
Tissue preparation techniques, especially fixation methods, also influence the thickness and appearance of the BM structure.17 Studies using other techniques such as cryofixation have shown that the BM is a homogenous layer, not laminated, and the lamina lucida is an artefact caused by fixation.17 BMs show variable thickness, but have been measured at about 100 nm by EM; however, this could be an underestimation due to dehydration caused during the preparation procedure, as proved by atomic force microscopy that showed twofold higher measurements.18 The BM may also be absent entirely, especially in embryonic tissues.19
The molecular components of BMs are classified into major and minor. Major structures are collagen IV, laminin, nidogens and perlecan while minor components include fibronectin and collagens XV and XVIII.15 20 Collagen IV is the main constituent representing approximately 50% of the BM.15 21 It is a non-fibrillar collagen which presents in a network with different six alpha chains known as α1 (IV) through α6 (IV).22 The chains α1 (IV) and α2 (IV) are known as the classical chains and are represented in BMs from all tissue types, whereas the other chains show variable distribution throughout differing tissues. The α5 (IV) and α6 (IV) chains are present in breast tissue.23–25 Laminins are a family of large glycoproteins consisting of α, β and γ chains. There are 15 different heterotrimeric isoforms of laminin resulting from 12 genetically different types of these chains. Laminin associates with other major structures of the BM to form the basic architecture of BMs and plays a key role in cell attachment and differentiation.20 26 Both laminin and collagen IV components are present diffusely along the basal lamina of the BM but are not localised to particular regions. Nidogen is a glycoprotein existing in two forms, N1 and N2, which bridge with collagen IV and laminin for BM stability.15 27 28 Perlecan is a heparan sulfate proteoglycan (HSPG) which contains binding sites for collagen IV and laminin, forming a bridging network and acting as a growth factor reservoir which affects adhesion and migration.15 29 30
The minor components of the BM are variable according to the tissue, and their functions and clinical significance remain unclear. Fibronectin is a glycoprotein present in lamina reticularis linking it with the basal lamina enhancing structural integrity. Collagen XV plays a role in BM maintenance, while collagen XVII is a non-fibrillar collagen binding with HSPGs which supports cell proliferation and migration. Collagens XV and XVIII are responsible for interconnecting the BM with the underlying connective tissue.31 There are also other minor components such as secreted protein acidic and rich in cysteine (SPARC) and fibulins.15 30 32
BM staining techniques and its pitfalls
H&E stain, which is the gold standard stain pathologists use routinely in practice, can highlight the BM around the ductolobular system and around proliferating ducts, but it has limitations as some of BM structures do not stain well.33 Periodic acid-Schiff (PAS) is a special stain used for BM detection which is demonstrated as a magenta line lying under the epithelium. PAS staining is based on detection of glycogen and mucus, staining mucus or glycogen producing tissues stronger than the collagen-rich BM. When diastase is added to break down glycogen (PAS/D), the BM appears as a pink line.33 BMs also contain reticular fibres, which appear as black lines when silver-based stains are applied. Silver stains show greater levels of contrast than PAS stains as peridot oxidation results in aldehydes which are detected as black colloidal silver deposits following application of silver nitrate solution.34 A comparative study of the various BM stains showed that PAS and silver stains produce the strongest reactions and are therefore arguably better than other connective tissue stains including Masson’s trichrome, Mallory stain and Wilder’s reticulum stain.35 Another study compared H&E, PAS and acriflavine (fluorescent periodic acid) stains showing that BM continuity, contrast and pattern are better determined by acriflavine than PAS as it showed the fibrillar pattern of the BM in which reticular fibres appeared black while collagen fibres appeared golden brown.33 IHC, alongside antibodies against the most ubiquitous BM components collagen IV and laminin, is now used more frequently in routine practice for BM identification.36 However, there are some limitations as cross-reactivity, propensity to stain vasculature structure37 and the expression of some of these markers in the malignant epithelial cells9 can complicate interpretation.
BM in normal breast tissue
Mammary gland cellular anatomy varies through different stages of development including puberty, pregnancy, lactation and menopause.6 During puberty, the terminal buds develop at the tip of the ducts. The BM at the bulbous tip of the terminal bud is thin (around 104 nm) and it becomes thicker at the bud neck region reaching 1.4 μm.38
EM was used in the past to detect BMs but it is technically challenging for routine use in breast pathology.39 When visualised using H&E staining and light microscopy, BMs appear as a fine line at the periphery of the normal parenchymal elements of the breast (TDLUs and ducts). However, this is often difficult to demonstrate and special stains such as PAS are often used to identify BMs. Despite the availability of special stains and IHC, the detection of BMs in breast lesions remains technically challenging, especially given the reduction in people who are experienced in interpretation of the staining findings. Therefore, the use of MEC markers has largely replaced the use of BM stains in routine practice. However, some pathologists still rely on BM markers and its diagnostic use remains valuable in certain situations where MEC markers provide limited value such as MGA.12
The BM structure and composition are different in normal, reactive hyperplastic and neoplastic benign and malignant breast lesions.40 Some malignant breast lesions secrete BM-like material in the extracellular spaces in the breast41 42 and in the lymph nodes,43 and some tumours express some of the BM components in the malignant epithelial cells and these should be distinguished from the native BM that denotes the in situ nature of breast lesions. This is observed more in tumours showing basal/MEC phenotypes including some salivary gland-like tumours. However, some conventional-type ductal no-special-type (NST) carcinomas have a BM-like structure around the invasive foci in the breast or in metastatic tumours particularly around DCIS-like structures in the nodes (revertant DCIS).44
Comparisons between BM and its mimickers
BMs share many chemical and ultrastructural features with the ECM in the interstitial stromal tissue36 and with some reactive processes such as scar tissue and pseudocapsule. In addition, BM-like material is secreted by some invasive breast tumours (table 1).
Characteristics of the breast basement membrane (BM) and its mimickers
The components of the BM are secreted by many types of cells in both normal and malignant conditions, and these can help differentiate the native BM secreted by MECs from its histological mimickers (table 2).
Cells secrete basement membrane (BM) components
While the non-fibrillar collagen IV is the most abundant component in the BM, fibrillar collagens I, III and V are the major components of the ECM.45 The ECM contains some collagens that act as anchoring fibrils (VII, VI, XXVI and XXVIII), which play a key role in interconnecting matrix components and separate structural components within tissues. Chondroitin sulfate and glycoprotein fibronectin, which is a minor component of the BM, are major components of the interstitial ECM and have a bridging role in organising matrix and cell–matrix interactions.46 Interstitial ECM contains matricellular proteins which are glycoproteins that can interact with matrix components, growth factors and cell surface receptors which represent a regulatory function in matrix–cell communications.47 48
Two main families of PGs are represented in interstitial ECM, namely hyalectans and small leucine-rich PGs, which have a role in matrix organisation, cell signalling, inflammation processes and collagen fibrillogenesis. Perlecan and agrin are types of PGs which are found in BMs, and they are responsible for interactions with matrix components.31 Unlike in BMs, elastin is a major component in ECM and accumulates as microfibrils composed of fibulins and fibrillin. These microfibrils insert into the BM binding to perlecan anchoring the BM to interstitial ECM.31 49
The interstitial ECM contains several key proteins which can be demonstrated by IHC and are different from those in the BM. These include collagen I, laminin 1, thrombospondin 1, tenascin C, fibulin 1 and α-smooth muscle actin (α-SMA),50 51 in addition to the stromal cell markers such as CD73, CD90 and CD105.52 However, collagen IV and laminin are the main markers for BM, and the latter is expressed in stromal cells.53 54 Other proteins, such as SPARC and fibronectin, are also expressed in the BM,55 and α-SMA is one of the markers for MECs.7 Some collagens known as anchoring fibrils are found in the BM zone and have been used recently to demonstrate BMs; these include collagens XV, XIX56 and VII.57 The stroma of the breast also contains adipocytes, fibroblasts, blood vessels and nerves. The blood vessels have their own laminin-rich BM. The stroma communicates with the epithelium despite their separation by the BM, and stromal changes are observed in the early stages of malignancy.36
Reactive fibrotic stroma is defined as a new microenvironment due to stromal reactions in response to external stimuli, such as injury or cancer, results in ECM remodelling, inflammatory cells, angiogenesis, growth factor release and desmoplastic reactions.58 If such reactive stromal elements are seen at the periphery of expansile breast lesions it can result in a band-like collagenised tissue mimicking thickened BM or a capsule (pseudocapsule). Quantitative analysis of ECM components in scar tissue showed more collagen I than ECM, with thicker less organised bundles, instead of exhibiting a basket wave appearance, with decreased elastin and fourfold increases in glycosaminoglycans.59 Histologically, scar tissue has a high mesenchymal density accompanied by high vascularity.60 Similar to the BM, scar tissue is negative for CD34,61 which is positive in normal intralobular mammary stroma.62 The α-SMA marker remains negative in scar tissue63 and normal stroma64 while it is positive in reactive stroma.65 Unlike native BMs, scar tissue and ECM show a negative reaction following silver staining,60 as does the reactive stroma in tumours.66 Components of BM examined in scar tissue revealed a BM-like structure with negative expression of collagen IV and positive laminin expression.60 67
It is also important to note that the various biochemical components of the BM can be detected in cancer cells and previous studies have linked such expression to tumour behaviour.9 10 36 Proteomic analysis of breast cancers (BC) has shown that only stromal cells secrete laminin and collagen IV in poorly metastatic tumours; however, in highly metastatic cancers, both stromal cells and tumour cells produce these BM proteins.68 Tumour-associated BM materials and their structures are likely to be different from the native BM present around normal parenchymal structure in the breast and around benign and in situ lesions,69 in structure, arrangement and anatomical localisation and in terms of cell origin and functions. Despite this knowledge, data relating to the nature and functions of extracellular BM materials associated with invasive tumour foci remain to be elucidated.
Diagnostic use of the BM in breast pathology
The basic and original definition of invasive breast carcinoma is based on BM breaching with stromal infiltration, involving the migration of invasive tumour cells in the stroma with potential local and distant spread.7 While the diagnostic use of BM is still valid, and some pathologists rely on BMs to distinguish in situ from invasive tumours at least in certain lesions,7 routine assessment of BM in practice is often challenging.70 Here we consider the challenges involved in diagnostic applications of BM assessment in breast pathology.
BM in DCIS
In situ carcinoma associated with microinvasion shows focal fragmentation and thinning, and disruption of the BM typically at the site of the microinvasion.71 This may support the use of BMs to differentiate in situ from invasive tumours in the breast. Chen and colleagues studied the BM using multiphoton microscopy and reported that the BM is intact but enlarged compared with normal BM in atypical ductal hyperplasia (ADH), while the BM is similar to ADH with straighter collagen fibres resulting from duct expansion in low-grade DCIS. In contrast, high-grade DCIS presents with varying collagen, thin in some areas and dense in others, with some ducts displaying disrupted BMs with microinvasion.72 Despite the differences in the structure, composition and arrangement of the BM components in DCIS compared with native BMs surrounding normal ductolobular system, it is believed that the DCIS BM is a modification of the native BM rather than a new structure. Therefore, their existence denotes the intraductal/intralobular nature of the proliferating epithelial cell components (figure 2).
Ductal carcinoma in situ photomicrographs. (A) H&E-stained tissue with a thickened basement membrane (BM) surrounding the gland. (B) Picaro Sirius Red (PSR)-stained tissue showing a thickened BM surrounding the gland. Magnification ×20. Scale bar=100 µm.
Some cases of high-nuclear-grade DCIS have exhibited thickened BMs together with inflammation and fibrosis. This has resulted in the perception that the well-developed layer of fibrous tissue (pseudocapsule) at the epithelial stroma interface around EPC may represent native BMs characteristic of in situ lesions.11 73 74 However, there are several lines of evidence to indicate that this layer likely represents florid and abnormal proliferation of the BM-like material secreted either by the proliferating malignant cells or the surrounding stromal cells.11 The presence of similar peripheral pseudocapsule around EPC at metastatic and recurrent sites outside the breast supports that such a structure is a reactive process initiated by the tumour itself73 74 (figure 3). This likely reflects the interactions between such a slowly growing EPC with expansile growth and pushing margins and the surrounding stroma, perhaps exaggerated by the cystic nature of EPC with leakage of cyst content into the stroma and a secondary stromal reaction. It is not infrequent to find stroma reactive changes similar to that seen at the previous biopsy site around benign and malignant intraductal papillary lesions across multiple areas, excluding the possibility of biopsy site-related changes.11 73 74

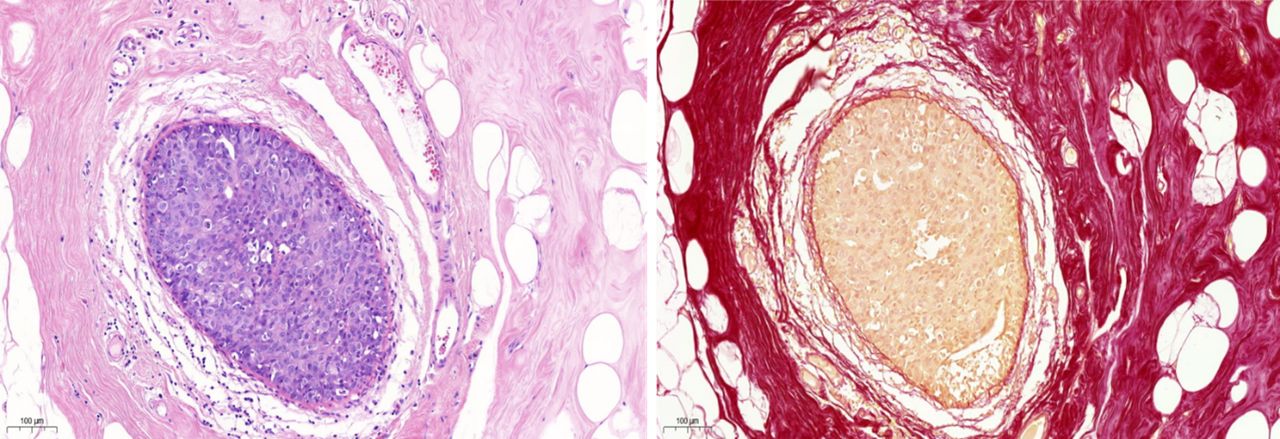
Encapsulated papillary carcinoma photomicrographs. (A) H&E-stained tissue showing thick fibrous capsule surrounds the gland. (B) Picaro Sirius Red (PSR)-stained tissue showing a thick fibrous capsule, red in colour, surrounding the epithelial cells stained yellow. Magnification ×20. Scale bar=100 µm.
Studies have shown that EPC lesions express higher levels of transforming growth factor beta 1 compared with conventional-type DCIS and invasive carcinomas, which plays a role in the development of the thick fibrous capsule, supporting the hypothesis that the EPC capsule is a reactive rather than a thickened expanded native BM resulting from distention by the proliferation of neoplastic papillary.74 Indeed, differentiation of a native BM from a reactive BM surrounding an invasive lesion is clinically important for early diagnosis and proper management of the patient, when considering implications of management in situ, and for benign versus invasive tumour diagnosis.
BM in other breast lesions
The BM is used more frequently when diagnosing MGA to differentiate it from tubular carcinoma. MGA is a clonal neoplastic proliferative lesion that shows infiltrating rounded glands surrounded by a well-developed BM.75 MGA lacks peripheral MECs and has an infiltrative appearance, so it mimics invasive breast lesions. In this context, MEC markers do not help differentiate MGA from tubular or other low-grade NST carcinomas with prominent tubule formation. In addition, due to the lack of the desmoplastic stromal reaction and the characteristic immunoprofile (oestrogen receptor negative and strong S100 positivity), BM markers are helpful, and they are frequently used in routine practice to distinguish MGA from invasive carcinoma with prominent tubule formation. Perhaps this is currently the main diagnostic use of BM markers in the breast in routine practice due to ease of interpretation with the thickened BM layer around the tubules of MGA with an absence of BM structures in tubular carcinoma. Despite this, experience in undertaking interpretation of BM staining pattern is required and the role of the BM in differentiating atypical MGA from foci of acinic cell carcinoma (AC) is less well defined. The BM surrounding the glands of MGA can be identified with PAS/D-PAS, collagen IV and laminin immunostaining.69 The characteristic infiltrative nature and the growth patterns of MGA suggest that the BM layer is not native and is reproduced by the neoplastic cells.33 This is an area for further research and a comparison of the ultrastructure and composition between MGA BM and native BM is warranted.
Lactational changes and apocrine metaplasia are two benign metaplastic changes in the breast that may show reduced or focal absence of MEC layers. Apocrine metaplasia can also exhibit cytological atypia (apocrine atypia) and lactational changes may also present with nuclear enlargement and irregularity raising suspicion of invasive carcinoma in cases with reduced or absent MECs. The use of BM markers can confirm the benign in situ nature of these lesions. Moreover, the BM has an important role in differentiating sclerosing adenosis from tubular carcinoma as IHC studies showed that continuous BMs surround the tubules of sclerosing adenosis despite perineural invasion in some cases; however, BMs are absent in tubular carcinoma.40 Collagenous spherulosis is a benign lesion characterised by multiple cysts composed of collagenous material surrounded by proliferative MECs and epithelial cells. It is associated with benign breast lesions and may be misdiagnosed as an adenoid cystic carcinoma (ACC). Studies have suggested that these spherules may represent BM material.76 Collagenous spherulosis exhibits positive staining for collagen IV and laminin; however, some showed that it contains only basal lamina which is duplicated.77
ACC is a rare breast carcinoma and is considered as a variant of the triple negative BC molecular subtype (figure 4); however, it has a better prognosis. ACC presents with a biphasic morphological pattern and the glands are surrounded by BM-like material.78 79 IHC staining of fibronectin and laminin within ACC demonstrate pseudocystic spaces containing disturbed BM-like material, mainly fibronectin and laminin that are lined by neoplastic cells. Both proteins were observed around the entire peripheral region of neoplastic cell clusters.80 Ultrastructural EM studies showed ACCs were lined by an uninterrupted BM consisting of lamina lucida and lamina densa which were characterised by their extraordinary thicknesses.81 These pseudocysts were surrounded by neoplastic cells, and as a result it was obvious that the BM lamina densa was produced by the tumour cells.81 The BM-like material within the pseudocyst contains the molecules of BM structures produced by the surrounding tumour cells which have basal/MEC characteristics. The presence of occasional vascular capillaries within these pseudocysts suggests invagination of the surrounding stroma into the tumour cribriform masses or solid structures. However, the ultrastructural features and the composition of the material in the pseudocysts are consistent with that of the BM and not that of the interstitial ECM stroma which support tumour secretion by cells having basal/MEC differentiation, in a way mimicking the development of the native BM in normal tissue. The indolent behaviour of ACCs may also reflect the degree of differentiation of these tumours which tries to recapitulate the normal tissue and secrete BM-like material. The correlation between anaplastic features and BM disintegration supports the role of the BM in the process of invasion.82 This may explain the better prognosis observed where the tumours have BM-like material.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
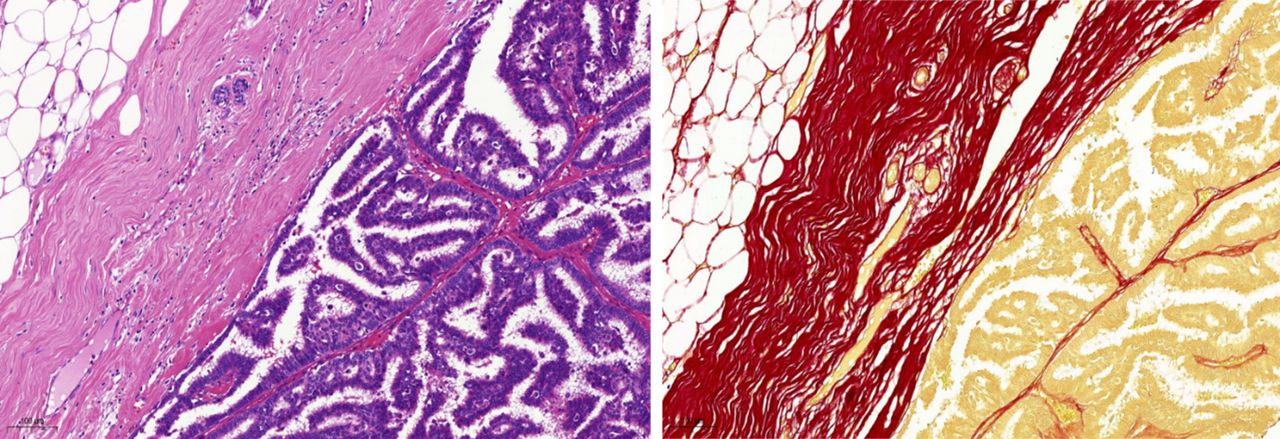
Adenoid cystic carcinoma tissue photomicrographs. (A) H&E stain showing clusters of epithelial cells in cribriform-like pattern surrounded by eosinophilic basement membrane (BM)-like material (horizontal arrows) and microcysts filled with solid spheres of BM material (vertical arrows). (B) Picaro Sirius Red (PSR) stain showing a cluster of epithelial cells in a cribriform pattern (yellowish colour) surrounded with red circular BM-like material (horizontal arrows) and the microcysts shown as dark red spheres of BM material (vertical arrows). Magnification ×40. Scale bar=20 µm.
AC is a rare type of salivary gland like breast carcinoma. AC with prominent well-differentiated acinar-like structures may resemble atypical MGA. Although both lack an MEC layer, AC also lacks a BM structure.83 Some cells in invasive breast carcinoma have shown intracytoplasmic staining of laminin.71 84–87 In addition, IHC studies have shown that BM components, especially HSPG, are synthesised by the rough endoplasmic reticulum in some BC cells88 (table 3).
Basement membrane (BM) and myoepithelial cells (MEC) in various breast lesions
In conclusion, the fundamental definition of cancer invasiveness is based on BM penetration; however, some invasive BCs exhibit BM-like material surrounding the neoplastic cells and some invasive tumours create a reactive capsule that may mimic native BMs. These structures are different in both composition and structure from the native BM that surrounds the ductolobular system of the breast. Knowledge of such differences between native BMs and mimicking tissues is likely to help provide more accurate interpretation of histological findings. Further studies are needed to provide better explanations about the BM changes in DCIS and the early invasive process and the relationship between the development of BM-like material in invasive lesions and tumour differentiation and behaviour.
Ethics statements
Patient consent for publication
Ethics approval
This work was approved by the Yorkshire and the Humber-Leeds East Research Ethics Committee (REC reference: 19/YH/0293) under the IRAS Project ID: 266925. Informed consent was obtained from all individuals prior to surgery to use their tissue materials in research. All samples were properly coded and anonymised. All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Declaration of Helsinki and its later amendments or comparable ethical standards. This article does not contain any studies with animals performed by any of the authors.
Acknowledgments
SFAG is supported and funded by the Egyptian Ministry of Higher Education and Scientific Research.
References
Footnotes
Handling editor Cheok Soon Lee.
Contributors SFAG and ER planned the review. SFAG wrote the manuscript. CR, CA, NM and ER reviewed and amended the manuscript.
Funding The authors have not declared a specific grant for this research from any funding agency in the public, commercial or not-for-profit sectors.
Competing interests None declared.
Provenance and peer review Not commissioned; internally peer reviewed.