Article Text

Abstract
Background Miscoding mutations of the TET2 gene, which encodes the α-ketoglutarate-dependent enzyme that catalyses the conversion of 5-methylcytosine to 5-hydroxymethylcytosine, thus producing DNA demethylation, have been detected in 10–25% of acute myeloid leukaemias lacking IDH1/2 mutations. Most low-grade diffuse gliomas carry IDH1/2 mutations (>85%), but molecular mechanisms of pathogenesis in those lacking IDH1/2 mutations remain to be elucidated.
Methods Miscoding mutations and promoter methylation of the TET2 gene were screened for in 29 low-grade diffuse gliomas lacking IDH1/2 mutations.
Results Single-strand conformational polymorphism followed by direct sequencing showed the absence of miscoding mutations in TET2. Methylation-specific PCR revealed methylation of the TET2 promoter in 5 of 35 cases (14%). In contrast, none of 38 low-grade diffuse gliomas with IDH1/2 mutations had TET2 promoter methylation (p=0.0216).
Conclusion Results suggest that TET2 promoter methylation, but not TET2 mutation, may be an alternative mechanism of pathogenesis in a small fraction of low-grade diffuse gliomas lacking IDH1/2 mutations.
- TET2
- IDH1
- IDH2
- glioma
- promoter methylation
- neuromuscular
- neuropathology
- neuro-oncology
- molecular oncology
- paediatric pathology
- brain tumourscancer research
- molecular pathology
Statistics from Altmetric.com
- TET2
- IDH1
- IDH2
- glioma
- promoter methylation
- neuromuscular
- neuropathology
- neuro-oncology
- molecular oncology
- paediatric pathology
- brain tumourscancer research
- molecular pathology
Introduction
IDH1/2 mutations are very frequent (>85% of cases) in astrocytic and oligodendroglial diffuse gliomas (WHO grades II and III) as well as in secondary glioblastomas (WHO grade IV) that are derived from diffuse astrocytomas or anaplastic astrocytomas.1 2 In contrast, IDH1 mutations are very rare or absent in other neoplasms of the central nervous system or tumours of other organ sites, including cancers of the bladder, breast, stomach, colorectum, lung, ovary and prostate.3–5 The only exceptions so far include acute myeloid leukaemia (AML; up to 20%),6–11 melanomas (approximately 10%)12 and anaplastic thyroid cancer (approximately 10%).13 In contrast to diffuse gliomas in which IDH1 mutations are significantly more frequent than IDH2 mutations,14 IDH1 and IDH2 mutations were detected at similar frequencies in AML.15 16
Recently, the TET2 (ten-eleven-translocation 2) gene at chromosome 4q24 has been found to be mutated in 10–25% of cases of AML, myelodysplastic syndrome and myeloproliferative neoplasms.15 17 Interestingly in AML, IDH1/2 mutations and TET2 mutations were mutually exclusive, and importantly, both IDH1/2 and TET2 mutations were associated with epigenetic defects and a hypermethylation signature.15 These observations suggest that TET2 mutations represent an alternative molecular mechanism in the development of AML lacking IDH1/2 mutations.
Epigenetic defects that resemble those seen in AML with either IDH1/2 or TET2 mutations have been recognised in gliomas carrying IDH1/2 mutations. In a study by Noushmehr et al,18 a distinct subset of glioblastomas showed concerted CpG island methylation at a large number of loci; these tumours typically carry IDH1/2 mutations. In a study of 131 brain tumours, hypermethylation of CpG loci was strongly associated with IDH1/2 mutations.19
These findings prompted us to screen for TET2 mutations in low-grade diffuse gliomas lacking IDH1/2 mutations. We also assessed TET2 promoter methylation, since this has recently been reported in a small fraction (4.4%) of myeloproliferative neoplasms (essential thrombocythemia).20
Materials and methods
Tumour samples
A total of 73 tumour samples of low-grade diffuse gliomas (WHO grade II) were obtained from the Department of Neuropathology, University Hospital Zürich, Switzerland, Department of Neuropathology, University Hospital Frankfurt, Germany, Institute of Neuropathology and Department of Neurosurgery, University Hospital Münster, Germany, Department of Neurosurgery, University Hospital Bern, Switzerland, and Institute of Neuroscience, Bordeaux, France.
Low-grade diffuse gliomas lacking IDH1/2 mutations (total 35 cases) were diffuse astrocytoma (17 cases), oligoastrocytoma (6 cases) or oligodendroglioma (12 cases); those carrying IDH1/2 mutations (total 38 cases) were diffuse astrocytoma (17 cases), oligoastrocytoma (9 cases) or oligodendroglioma (12 cases). The mean age of the patients was 40.1±14.7 years (range 5–82 years). Fifty-six patients were treated with surgery alone and 13 patients with surgery followed by radiotherapy. None of the patients were treated with chemotherapy. This study was approved by ethical committees in each collaborative centre as well as by the IARC ethical committee.
TET2 mutations
We screened for mutations of the TET2 gene at exons 3–11 containing the conserved domains 1 and 2, in which approximately 90% of all mutations have been detected in AML and myeloid disorders17 21 22 in 29 low-grade diffuse gliomas lacking IDH1/2 mutations. Single-strand conformational polymorphism (SSCP) analysis was carried out to pre-screen for mutations in exons 3–11 of the TET2 gene, as described previously.23 The primer sequences are available on request. DNA samples containing IDH1 mutations and H2O served as positive and negative controls, respectively. Samples exhibiting mobility shifts in SSCP analyses were subsequently analysed by direct sequencing using an ABI 3100 PRISM DNA sequencer (Applied Biosystems, Foster City, California, USA) with the Big Dye Terminator cycle sequencing kit (ABI PRISM, Applied Biosystems).
TET2 promoter methylation
Methylation-specific PCR was carried out in 35 low-grade diffuse gliomas without IDH1/2 mutations and 38 low-grade diffuse gliomas with IDH1/2 mutations, to assess TET2 promoter methylation using primers reported by Chim et al.20 For each methylation-specific PCR reaction, we included universal methylated DNA (Chemicon International, Temecula, California, USA) as positive control, and normal blood DNA as negative control.24 Direct sequencing confirmed bisulfite-modified DNA in the positive control. Quality controls for bisulfite conversion were performed for each reaction, as previously described.25
Results
No miscoding mutations of TET2 were detected in the 29 low-grade diffuse gliomas lacking IDH1/2 mutations that we analysed.
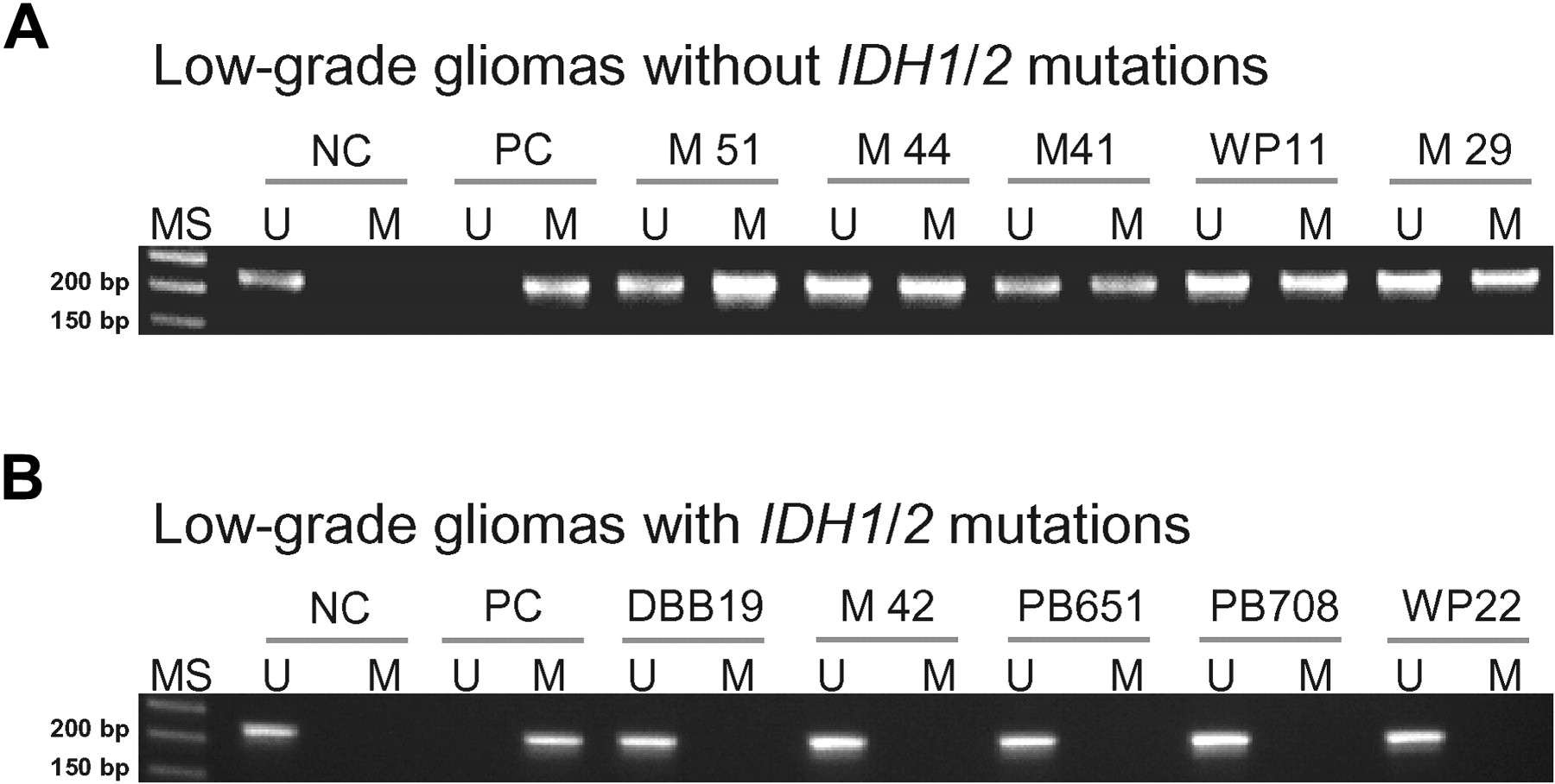
Methylation-specific PCR showed TET2 promoter methylation in 5 of 35 (14%) low-grade diffuse gliomas lacking IDH1/2 mutations (three diffuse astrocytomas and two oligodendrogliomas; figure 1). None of 38 low-grade diffuse gliomas carrying IDH1/2 mutations showed TET2 promoter methylation. The difference in frequency of TET2 promoter methylation in tumours with and without TET2 promoter methylation was statistically significant (p=0.0216). The median survival of patients was not significantly different between cases with or without TET2 promoter methylation.
{kind=link}
Methylation-specific PCR analysis of the TET2 promoter region in low-grade diffuse gliomas. (A) Without IDH1/2 mutations (14%); (B) with IDH1/2 mutations (0%). MS, molecular size marker; U, PCR product amplified by unmethylated-specific primers; M, PCR product amplified by methylated-specific primers; NC, normal control; PC, positive control (universal methylated DNA).
Discussion
Mammalian DNA contains two modified cytosine bases, 5-methylcytosine (5mC) and 5-hydroxymethylcytosine (5hmC).26 Of these, 5mC is known to play a significant role in epigenetic modification involved in gene regulation, X-chromosome inactivation, genomic imprinting and cancer development.26 In contrast, attention has only recently been drawn to 5hmC, with the report that the TET family catalyses the conversion of 5mC to 5hmC,26 27 and that one of the enzymes in this family, TET2, is frequently mutated in myeloid neoplasms.15 17 The formation of 5hmC can lead to DNA demethylation, which may contribute to the dynamics of DNA methylation.26 The conversion of 5mC to 5hmC by TET2 is α-ketoglutarate (α-KG)-dependent.27 28
IDH1 and IDH2 are enzymes that catalyse the interconversion of isocitrate and α-KG.7 15 29 30 IDH1/2 mutations impair enzyme affinity and dominantly inhibit wild-type IDH1/2 activity through the formation of catalytically inactive heterodimers,31 which leads to down-regulation of α-KG and up-regulation of 2-hydroxyglutarate.15 32 Decrease in α-KG expression results in up-regulation of HIF-1α and its targets, such as GLUT1, VEGF and PGK1 in U87MG malignant glioma cells.31 Thus, the functions of TET2 and IDH1/2 may be linked via α-KG.
The present study shows the absence of TET2 mutations in low-grade diffuse gliomas lacking IDH1/2 mutations. Thus, in contrast to AMLs, mutational inactivation of the TET2 gene is not a mechanism of pathogenesis in low-grade diffuse gliomas.
Recent studies suggest that TET2 is a tumour suppressor gene, since TET2 mutations have been detected in 10–25% of AMLs, myelodysplastic syndrome and myeloproliferative neoplasms,15 17 and TET2 promoter methylation has been reported in a small fraction (4.4%) of myeloproliferative neoplasms.20 Loss at the TET2 locus (4q24) has been observed in a small fraction of glioblastomas (2%).33 We postulated that TET2 promoter methylation may occur in gliomas. Indeed, in the present study we show TET2 promoter methylation in a fraction of low-grade diffuse gliomas lacking IDH1/2 mutations (14%). In contrast, none of 38 low-grade diffuse gliomas carrying IDH1/2 mutations had TET2 promoter methylation (p=0.0216). These results suggest that TET2 promoter methylation, but not TET2 mutation, may be an alternative mechanism of pathogenesis in a small fraction of low-grade diffuse gliomas lacking IDH1/2 mutations. However, the low frequency of methylation of the TET2 promoter in low-grade diffuse gliomas suggests that there may be other, as yet unknown, mechanisms of glioma development that do not involve the TET2 or IDH1/2 pathways.
Take-home messages
In contrast to acute myeloid leukaemia, ten-eleven-translocation 2 (TET2) mutations are absent in low-grade diffuse gliomas lacking IDH1/2 mutations.
TET2 promoter methylation is present in a small fraction of low-grade diffuse gliomas lacking IDH1/2 mutations.
TET2 promoter methylation, but not TET2 mutation, may be an alternative mechanism of pathogenesis in a small fraction of low-grade diffuse gliomas lacking IDH1/2 mutations
References
Footnotes
Funding DP is supported by the Fritz-Thyssen Foundation.
Competing interests None.
Ethics approval Ethics approval was provided by IARC.
Provenance and peer review Not commissioned; externally peer reviewed.