Article Text

Abstract
Objective: To examine the effects of thrombin on RANTES mRNA expression through protease activated receptor in synovial fibroblasts in patients with rheumatoid arthritis (RA).
Methods: A semiquantitative reverse transcriptase-polymerase chain reaction and reporter gene assay were performed using cultured human synovial fibroblasts from patients with RA. The up regulatory effects of thrombin on RANTES mRNA expression were tested. In addition, the roles of protease activated receptors (PARs) were analysed.
Results: PAR-1 and PAR-3, but not PAR-4, were expressed in synovial fibroblasts. Thrombin induced RANTES mRNA expression in a time dependent manner in synovial fibroblasts expressing PAR-1. A reporter gene assay showed that thrombin-induced RANTES gene expression was through PAR-1, but not PAR-3.
Conclusions: Thrombin induced RANTES mRNA expression through a PAR-1 mediated pathway, possibly indicating that thrombin has an important role in migration of inflammatory cells by RANTES to the synovium in patients with RA.
- rheumatoid arthritis
- RANTES
- thrombin
- protease activated receptor
- FCS, fetal calf serum
- GAPDH, glyceraldehyde-3-phosphate dehydrogenase
- PARs, protease activated receptors
- RA, rheumatoid arthritis
- RANTES, regulated upon activation, normal T cells expressed and secreted
- RT-PCR, reverse transcriptase-polymerase chain reaction
Statistics from Altmetric.com
- FCS, fetal calf serum
- GAPDH, glyceraldehyde-3-phosphate dehydrogenase
- PARs, protease activated receptors
- RA, rheumatoid arthritis
- RANTES, regulated upon activation, normal T cells expressed and secreted
- RT-PCR, reverse transcriptase-polymerase chain reaction
RANTES, a chemokine which causes the migration of memory type CD4+ T cells, is increased in the serum and synovial fluid of patients with active rheumatoid arthritis (RA).1 Ellingsen et al reported that synovial fluid levels of RANTES in patients with RA correlated positively with the migration of mononuclear cells towards synovial fluid.2 Furthermore, high levels of serum RANTES during the active stage of RA were significantly lowered by treatment with methotrexate.3 Additionally, in Lewis rats with adjuvant-induced arthritis, antibodies against RANTES are reported to ameliorate symptoms.4 Therefore, RANTES plays an important part in RA pathogenesis.
In contrast, coagulation abnormalities have also been shown to be a feature of RA. For example, it has been reported that clotting factors and fibrinolytic products, such as thrombin and D-dimer, are increased in synovial fluids of patients with RA.5 We have also reported that thrombin stimulated synovial cell proliferation through a protein kinase C mediated pathway.6 Recently, Marty et al showed that hirudin, a thrombin inhibitor, could both prevent onset of collagen-induced arthritis in mice and ameliorate established arthritis.7 Thrombin is a serine protease that cleaves fibrinogen to form fibrin monomers and uniquely cleaves cell surface receptors, known as protease activated receptors (PARs). Four members have recently been cloned and designated PAR-1, PAR-2, PAR-3, and PAR-4. Three of these members, PAR-1, PAR-3, and PAR-4, are cleaved by thrombin, whereas PAR-2 is cleaved by trypsin. PAR-1, especially, is known to be expressed in inflamed rheumatoid synovial tissues.8 However, little is known about whether PAR-3 and PAR-4 are expressed in synovial cells and whether thrombin induces RANTES expression.
In this study we found for the first time that thrombin increased RANTES mRNA expression through PAR-1, but not PAR-3, in synovial fibroblasts. These results suggest that thrombin may have an important role in the migration of inflammatory cells by RANTES to the synovium expressed PAR-1 in patients with RA.
MATERIALS AND METHODS
Cell cultures
Synovial tissue samples were obtained during orthopaedic surgery from the knees of eight patients with RA. The preparation of synovial fibroblasts was described previously.6 Suspended HeLa cells were kept in Joklik modified minimum essential medium (Gibco BRL) with 1% non-essential amino acids, 100 μg/ml penicillin, 100 U/ml streptomycin, and 10% fetal calf serum (FCS).
Reagents and plasmid constructs
Thrombin, a kind gift of Sankyo Co, Tokyo, Japan, was dissolved in distilled water. Human PAR-1 and PAR-3 cDNA were kind gifts from Dr Shaun R Coughlin (University of California, San Francisco). For PAR-1 and PAR-3 expression, FLAG-tagged pcDNA1PAR-1 and FLAG-tagged pcDNA1PAR-3 were generated from pBluescript SK-PAR-1 and pBluescript II SK-PAR-3 by polymerase chain reaction (PCR), respectively, and cloned into the BamHI site of pcDNA1, which contained a FLAG epitope inserted into the HindIII and BamHI sites of pcDNA1. RANTES promoter-luciferase reporter plasmid was a kind gift from Dr Alan M Krensky (Stanford University School of Medicine).
RNA extraction and semiquantitative reverse transcriptase-polymerase chain reaction (RT-PCR) analysis
Total RNA was extracted from cultured synovial fibroblasts as described previously.6 Synthesis of the first strand of cDNA and PCR analysis were performed according to instructions delivered with the RNA PCR kit (AMV) Ver.2 (TaKaRa) as described previously.6 For RANTES mRNA, samples from synovial fibroblasts were amplified at 94°C for five minutes, at 56°C for 90 seconds, and at 72°C for 120 seconds, followed by 28 cycles at 94°C for 30 seconds, at 56°C for 90 seconds, and at 72°C for 120 seconds. For PAR-1, PAR-3, and PAR-4 mRNA, samples from synovial fibroblasts were amplified at 94°C for five minutes, at 55°C for 30 seconds, and at 72°C for 90 seconds, followed by 40 cycles at 94°C for 30 seconds, at 55°C for 30 seconds, and at 72°C for 90 seconds. The following primers were used: for glyceraldehyde-3-phosphate dehydrogenase (GAPDH), sense 5`-ACATCGCTCAGACACCATGG-3`, antisense 5`-GTAGTTGAGGTCAATGAAGGG-3`; for PAR-1, sense 5`-ATCCCATGCAGTCCCTCT-3`, antisense 5`-GAAGAC AGCAGAGAAGGC-3`; for PAR-3, sense 5`-TCCATCCTTTC ACCTACCGGG-3`, antisense 5`-TAGCAGTAGATGATAAGC ACA-3`; for PAR-4, sense 5`- TGGCACTGCCCCTGACACTGCA-3`, antisense 5`- CCCGTAGCACAGCAGCATGG-3`; for RANTES, sense 5`-GCTGTCATCCTCATTGCTAC-3`, antisense 5`-TCCATC CTAGCTCATCTCCA-3`.
Transfection and luciferase assays
Transfection of suspended HeLa cells was performed by electroporation with a Bio-Rad gene pulser as described previously.9 Suspended HeLa cells were co-transfected with PAR-1 or PAR-3 expression vector (10 μg) and RANTES promoter-reporter plasmids (10 μg). β-Galactosidase expression plasmid was used as an internal control of transfection efficacy. Luciferase activities were measured with a luminometer (Lumat LB9501; Berthold).
RESULTS
PAR-1 and PAR-3 mRNA, but not PAR-4 mRNA, were expressed in synovial fibroblasts
In synovial cells, the relative levels of PAR mRNA expression clearly differed (fig 1). PAR-1 mRNA was strongly expressed in RA1, RA4, and RA5 synovial fibroblasts. Additionally, weak expressions of PAR-1 mRNA were seen in RA3, RA6, and RA7 synovial fibroblasts. In contrast, PAR-3 mRNA was expressed in RA1, RA5, RA6, and RA7. We found little mRNA expression of PAR-4 in any of the synovial fibroblasts tested. Thus, different levels of PAR-1 and PAR-3, but not PAR-4, were expressed in the synovial fibroblasts of patients with RA.
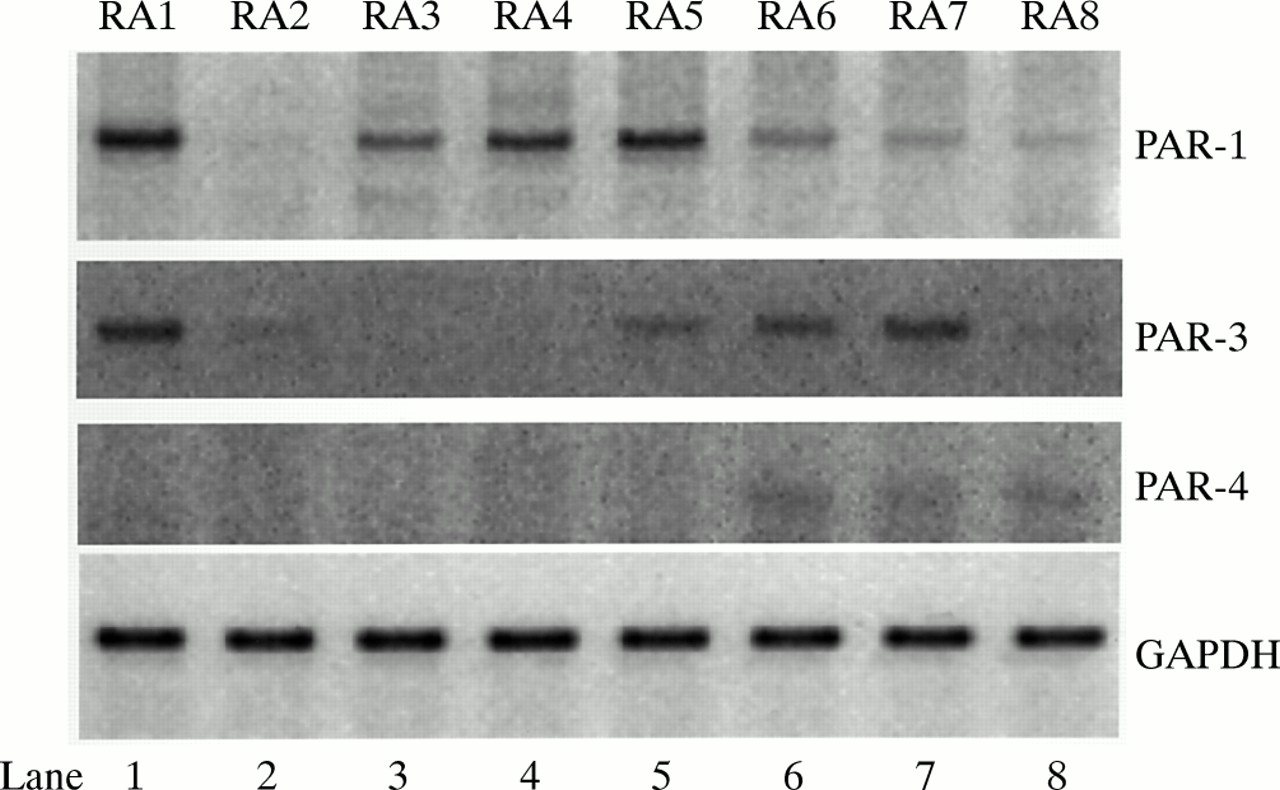
Expression of PAR-1, PAR-3, and PAR-4 mRNA in synovial fibroblasts from eight patients with RA (RA1–RA8).GAPDH was used as an internal control.
Thrombin-induced RANTES mRNA expression in synovial fibroblasts
To investigate the effects of thrombin on RANTES mRNA expression in synovial fibroblasts we used RA1, RA2, RA4, RA5, and RA7 synovial fibroblasts. Cells were starved for 24 hours in Opti-MEM (Gibco BRL, Grand Island, NY, USA) containing 0.5% FCS, then treated with 50 U/ml thrombin for the times indicated (fig 2). Cell viability was intact in the medium containing 50 U/ml thrombin for 48 hours (data not shown). Figure 2 shows that thrombin clearly induced RANTES mRNA expression in a time dependent manner in RA1, RA4, and RA5 synovial fibroblasts to a maximum level at 24 hours. In contrast, thrombin did not stimulate RANTES mRNA expression after 48 hours' incubation in RA2 and RA7 synovial fibroblasts. Therefore, thrombin up regulated RANTES mRNA expression in synovial fibroblasts expressing PAR-1, but not PAR-3.

Effect of thrombin on RANTES mRNA expression in synovial fibroblasts from patients with RA. A semiquantitative RT-PCR analysis was performed using total RNA in synovial fibroblasts from RA1, RA2, RA4, RA5, and RA7. After densitometric quantification, results were shown as a RANTES mRNA/GAPDH mRNA ratio. Results are the mean (SE) of three independent experiments.
Roles of PAR-1 and PAR-3 in thrombin-induced RANTES promoter activity
To test the effects of thrombin on RANTES gene expression, a luciferase assay was performed. A RANTES promoter-luciferase reporter plasmid and PAR-1 or PAR-3 expression vector were transiently co-transfected into a suspension of HeLa cells; we had verified that PAR-1 and PAR-3 mRNA were not expressed in suspended HeLa cells (data not shown). Thrombin increased RANTES gene expression, in a dose dependent manner, when PAR-1 was co-transfected with the reporter plasmid to suspended HeLa cells (fig 3, lanes 5–8). However, thrombin did not increase RANTES gene expression when PAR-1 was not transfected (fig 3, lanes 1–4). Moreover, thrombin did not influence RANTES gene expression in a PAR-3 transfected suspension of HeLa cells (fig 3, lanes 9–12). In addition, levels of PAR-1 and PAR-3 expression on suspended HeLa cells after transfection were almost equal (data not shown). Thus, thrombin induced RANTES expression at the transcriptional level through PAR-1, but not PAR-3.

{kind=link}
{kind=link}
{kind=link}
Effects of PAR-1 and PAR-3 on thrombin-induced RANTES gene expression. After transfection, cells were incubated with indicated concentrations of thrombin. After 24 hours, cellular extracts were prepared for a luciferase enzyme assay. Experiments were performed in quadruplicate. Levels of luciferase activity in unstimulated cells transfecting the reporter plasmid alone were taken as 1.0 (lane 1). Results are the mean (SE) of three independent experiments.
DISCUSSION
In this study we found that thrombin induced RANTES mRNA expression through PAR-1, but not PAR-3, in synovial fibroblasts from patients with RA. O'Brien et al showed that PAR-1 is the predominant thrombin receptor expressed in human endothelial cells and that despite the presence of PAR-3 mRNA in human endothelial cells, PAR-3 is either not expressed on the cell surface or is unable to support a thrombin response on its own.10 Our results also demonstrated that in human synovial fibroblasts PAR-1 and PAR-3 mRNA, but not PAR-4 mRNA are present. Furthermore, a PAR-1 mediated pathway is required for thrombin-induced RANTES gene expression in human synovial fibroblasts. Thus, PAR-1 may be the main receptor for thrombin in synovial fibroblasts as well as endothelial cells. However, a further transcriptional study should examine whether RANTES gene expression is induced by thrombin in synovial fibroblasts. In contrast with PAR-1, the role of PAR-3 in synovial fibroblasts is not at all clear. Mouse PAR-3 does not by itself mediate transmembrane signalling, suggesting that mouse PAR-3 functions as a cofactor for PAR-4 activation by thrombin.11 However, human PAR-3 function has remained poorly defined in the majority of cell types in which it has been identified.11
We further found that the levels of mRNA expression of PARs in synovial fibroblasts were clearly different among the patients with RA tested. Morris et al noted that cells expressing PAR-1 were present in rheumatoid synovia.8 In contrast, both osteoarthritic and normal synovia contained very few cells expressing receptor.8 Sequence analysis of the 5` flanking region of both PAR-1 and PAR-3 showed that neither a TATA box nor CCAAT sequences were evident, although nucleic acid motifs potentially involved in transcriptional gene regulation were present12; GATA motifs, octamer enhancer sequences, AP2-like sites, and Sp1 sites were included.12 With regard to PAR-1, Wu et al indicated that Sp1 transactivated promoter activity of PAR-1 and Sp3 inhibited Sp1 mediated activation.13 Moreover, thrombin itself significantly induced PAR-1 mRNA.7,14 In contrast, tumour necrosis factor α decreased the expression of PAR-1 in endothelial cells.15 Therefore, possibly, genetic and environmental factors regulate PAR-1 expression. However, regulation of the expression of PARs is not fully understood. In the future, we also need to examine the mechanism of expression of PARs in patients with RA.
In summary, we suggest that thrombin has a pivotal role in RA pathogenesis through RANTES expression. Furthermore, we have shown that activation of PAR-1 is required for thrombin-induced RANTES expression.
Acknowledgments
We thank Dr Shaun R Coughlin (University of California, San Francisco) and Dr Alan M Krensky (Stanford University School of Medicine) for providing the plasmid constructs.