Article Text

Abstract
Objective: To determine whether anti-Ro52 is an independent serum marker in connective tissue disease.
Methods: Over a two year period, 1727 consecutive antinuclear antibody (ANA) positive serum samples were analysed in parallel by double immunodiffusion with thymus/spleen nuclear extract and by line immunoassay with recombinant Ro52, recombinant La/SSB, and natural Ro60. Sera that were only reactive towards Ro52 were further analysed by a variety of additional anti-SSA/Ro detection methods and by specific anti-Ro52 and anti-Ro60 assays. Natural purified SSA/Ro was analysed by immunoblot and protein sequencing.
Results: Analysis of natural purified SSA/Ro (Immunovision, Springdale, AR) showed only Ro60 and no immunoreactive Ro52. Consequently, assays based on this substrate only identify sera with anti-Ro60 reactivity. Twenty serum samples showed anti-Ro52 without anti-Ro60 and anti-SSB/La on line immunoassay. By additional testing, 2/20 sera were found positive for anti-Ro60 reactivity. The remaining 18 sera were not identified by any of the classical anti-SSA/Ro assays and were considered to be reactive only with Ro52 and not with Ro60. This anti-Ro52 reactivity was confirmed by natural and recombinant Ro52 in 16/18 cases. 12/18 sera corresponded to connective tissue diseases.
Conclusion: Anti-Ro52 positive sera without any evidence of anti-Ro60 and anti-La/SSB reactivity can be considered as an independent group that is systematically missed by classical anti-SSA/Ro detection methods owing to a bias towards anti-Ro60 reactivity. The anti-Ro52 sera are precipitin negative, not retrieved by SSA/Ro enzyme linked immunosorbent assays (ELISAs) based on natural SSA/Ro, and show no specific ANA fluorescence staining pattern. These findings together with the clinical data indicate that anti-Ro52 should be considered as an additional and independent serum marker.
- anti-Ro52
- anti-SSA/Ro
- connective tissue disease
- ANA, antinuclear antibodies
- DID, double immunodiffusion
- ELISA, enzyme linked immunosorbent assay
- HPLC, high performance liquid chromatography
- mAb, monoclonal antibody
- PAGE, polyacrylamide gel electrophoresis
- PBS, phosphate buffered saline
- SDS, sodium dodecyl sulphate
Statistics from Altmetric.com
- ANA, antinuclear antibodies
- DID, double immunodiffusion
- ELISA, enzyme linked immunosorbent assay
- HPLC, high performance liquid chromatography
- mAb, monoclonal antibody
- PAGE, polyacrylamide gel electrophoresis
- PBS, phosphate buffered saline
- SDS, sodium dodecyl sulphate
Anti-SSA/Ro antibodies are the most frequently identified antinuclear antibodies (ANA).1 The major antigen bound by anti-SSA/Ro is the Ro60 protein associated with one of several hYRNAs. The SSB/La antigen is also associated with this Ro-ribonucleoprotein complex. More than 80% of anti-Ro sera defined by double immunodiffusion (DID) recognise a 52 kDa protein (Ro52) in immunoblot. The physical relationship between Ro52 and the Ro-ribonucleoproteins is controversial.2
To detect anti-Ro autoantibodies, laboratories can use Ouchterlony immunodiffusion or a purified natural SSA/Ro enzyme linked immunosorbent assay (ELISA).
We analysed consecutive ANA positive serum samples for monospecific anti-Ro52 reactivity. Eighteen samples were collected. These sera are missed by conventional anti-SSA/Ro tests because these detect only anti-Ro60. This and the clinical relevance of “monospecific” anti-Ro52 make anti-Ro52 an additional independent serum marker.
PATIENTS AND METHODS
Patients and serum samples
Over two years we consecutively identified 1727 ANA positive sera in our routine laboratory tests (University Hospital Gent, Belgium). Each ANA positive serum was analysed by indirect immunofluorescence on Crithidia luciliae (Immunoconcepts, Sacramento, CA), by DID using thymus/spleen nuclear extract (mammalian extracted nuclear antigen; Immunoconcepts, Sacramento, CA) and by ANA line immunoassay. Diagnoses were made according to classification criteria.3–5
Indirect immunofluorescence
ANA were determined on HEp-2 cells (Medica, Carlsbad, CA) at 1/40. Some sera were additionally analysed on HEp-2000 (Immunoconcepts). The result in the latter assay was reported Ro60 positive if the characteristic staining was present.6,7Crithidia luciliae was used for anti-dsDNA antibody detection.
Line immunoassay
Anti-Ro52 was detected by line immunoassay (INNO-LIA ANA K1090, Innogenetics NV, Zwijnaarde, Belgium). This assay contains Ro52, Ro60, La/SSB, SmB, SmD, Jo-1, RNP-A, RNP-C, RNP-70, Cenp-B, Scl-70, riboRNP, and histones. For Ro52, and La/SSB, the full size E coli derived recombinant proteins are applied, whereas for Ro60 the natural protein is used.
ELISA
The commercial Varelisa Ro antibodies (Pharmacia-Upjohn, Freiburg, Germany) (recSSA-ELISA) uses both baculovirus-expressed recombinant Ro52 and Ro60, coated in an unspecified ratio. Custom made ELISAs from Pharmacia Research and development laboratory are coated either with baculovirus-expressed recombinant Ro52 and Ro60 separately or with both proteins in a 50/50 ratio. The baculovirus-Ro52 coated ELISA was additionally incubated with reducing buffer.8 Also, an ELISA with natural SSA/Ro (ENA-LISA “SSA”, BMD, Brugge, Belgium) (nat SSA-ELISA) was used.
Immunoblot analysis
Immunoblotting of E coli-expressed recombinant Ro60 and natural SSA/Ro (bovine nature, Immunovision) was performed as described.9 Mouse IgG1 antihuman Ro52 and anti-Ro60 monoclonal antibodies (mAbs; Cappel, OT, Turnhout, Belgium) were detected with rabbit antimouse IgG (Sigma, St Louis, MO, USA). HeLa-S100 extract was separated on a 10% sodium dodecyl sulphate (SDS)-polyacrylamide gel with modified acrylamide/bisacrylamide ratio (172.4/1) to allow efficient separation of Ro52 and the La/SSB protein, and blotted onto nitrocellulose.10
Purifying antibodies from immunoblot with natural Ro60
Nitrocellulose blots with natural Ro60 were incubated with serum in blocking buffer overnight and washed with Tris buffer before elution of bound antibodies with elution buffer (0.1 M glycine-HCl pH 2.2; 500 mM NaCl, 0.5% Tween 20, 1% bovine serum albumin). Affinity purified antibodies were neutralised by 1 M Tris-HCl pH 8.45.
Double immunodiffusion
DID (Immunoconcepts) in agar was performed with thymus/spleen nuclear extract. Antibody specificity was determined by comparison with an anti-SSA/Ro reference serum. DID was also performed with natural SSA/Ro (Immunovision, Springdale, AR).
Protein sequencing of natural SSA/Ro (Immunovision)
Coomassie blue stained bands loaded with natural SSA/Ro were excised and underwent proteolytic digestion.11 The peptide mixture was separated by high performance liquid chromatography (HPLC). Manually recovered peptide fractions were then subjected to sequencing by Edman degradation using a Procise 492 equipped with an online 140C phenylthiohydantoin analyser (Perkin Elmer, Foster City, CA). Amino acids were identified by HPLC.
Protein immunoprecipitation
Protein A beads (Protein A-Sepharose 4 Fast Flow, Pharmacia Biotech) were prepared for protein immunoprecipitation by washing and blocking with RIPA buffer, followed by washing with phosphate buffered saline (PBS). After centrifugation and removal of the supernatants, the beads were resuspended in PBS. Cell lysate of 35S (Amersham Pharmacia) labelled HeLa cells (ECACC 85060701) was incubated with protein A beads to pre-clear the lysate. After centrifugation, the supernatant was incubated with serum and PBS to form antigen-antibody complexes, followed by incubation with protein A beads. After centrifugation the suspension was washed with RIPA buffer followed by washing with 62.5 mM Tris-HCl pH 6.8. Elution was performed by adding sample buffer and boiling at 95°C. After immunoprecipitation, bound 35S labelled HeLa cells were analysed by SDS-polyacrylamide gel electrophoresis (PAGE) on 8% Laemmli gel. After electrophoresis the gels were fixed, dried, and autoradiographed using Fuji Medical x ray film (ref 03G220 ) on Gevamatic 60 (Agfa-Gevaert).
RESULTS
Analysis of natural SSA/Ro
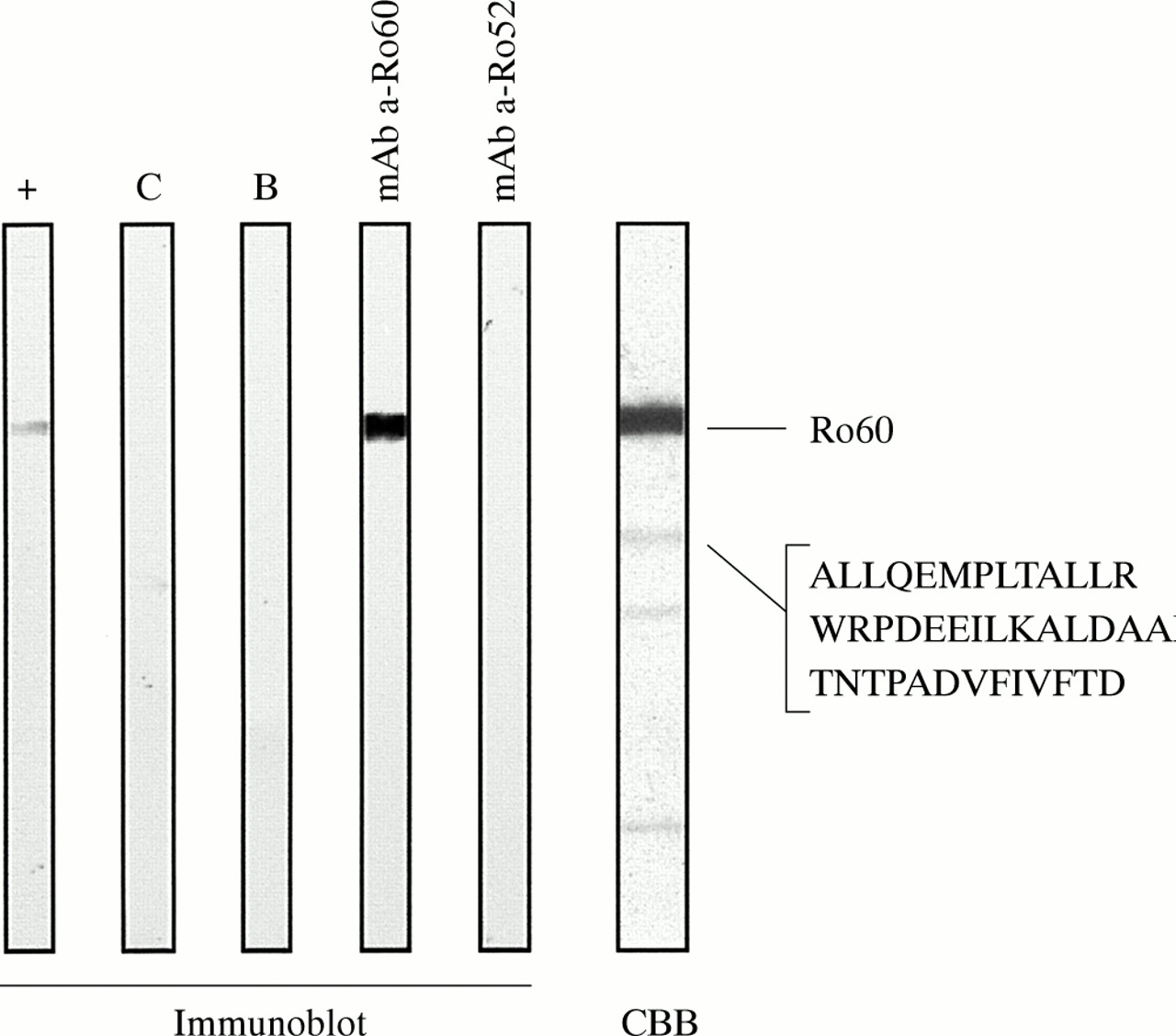
Analysis of natural SSA/Ro (Immunovision) Coomassie stained by SDS-PAGE showed one major band at 60 kDa while the closest minor visible band migrated at 42 kDa. Sequencing of this 42 kDa band disclosed three peptides (aa311-aa322; aa382-aa497; aa496-aa508) attributable to the Ro60 protein. Upon blotting the natural SSA/Ro, no reactivity was found using monoclonal anti-Ro52 antibodies. Monoclonal anti-Ro60 antibodies bound the major band. We conclude that the referred natural SSA/Ro source contained no immunoreactive Ro52 (fig 1).

Analysis of natural purified SSA/Ro (Immunovision). No Ro52 was detected on immunoblot using different reagents. Reference serum with anti-Ro60 and high titre of anti-Ro52 antibodies (lane +). Control serum negative for anti-SSA (lane C). Blank control (lane B). Monoclonal anti-Ro60 antibody (lane mAb a-Ro60). Monoclonal anti-Ro52 antibody (lane mAb a-Ro52). Coomassie brilliant blue staining of natural purified SSA/Ro and protein sequencing of the 42 kDa band shows three peptides attributable to Ro60 (lane CBB). Reactivity of the monoclonal anti-Ro52 antibody in western immunoblot was confirmed (see fig 2, lane mAb a-Ro52).
Anti-Ro52 without anti-Ro60
We consecutively identified 20 sera displaying anti-Ro52 reactivity without anti-Ro60 and without anti-SSB/La reactivity on line immunoassay. Anti-Ro60 reactivity was additionally analysed by immunoblot with natural and recombinant Ro60; by HEp-2000; by a recombinant and natural SSA-ELISA; and by DID with natural Ro60. When these tests were combined, anti-Ro60 was additionally found in two sera (Nos 6598, 10581) (table 1). The remaining 18 sera exclusively recognised Ro52 and not Ro60 and SSB. To ensure that they were not reactive with Ro60, we tested eight sera by protein immunoprecipitation to exclude conformation dependent-anti-Ro60 antibodies. None showed anti-Ro60 reactivity.
Analysis of anti-Ro52 sera without evidence of anti-Ro60 by different anti-Ro52 and anti-Ro60 methods. Values shown in bold exceed the cut off value
Confirmation of anti-Ro52
To confirm anti-Ro52 reactivity we analysed the sera by other anti-Ro52 detection methods: HeLa-S100 immunoblot (fig 2) accounting for natural Ro52, and ELISAs with baculovirus derived Ro52 (table 1).
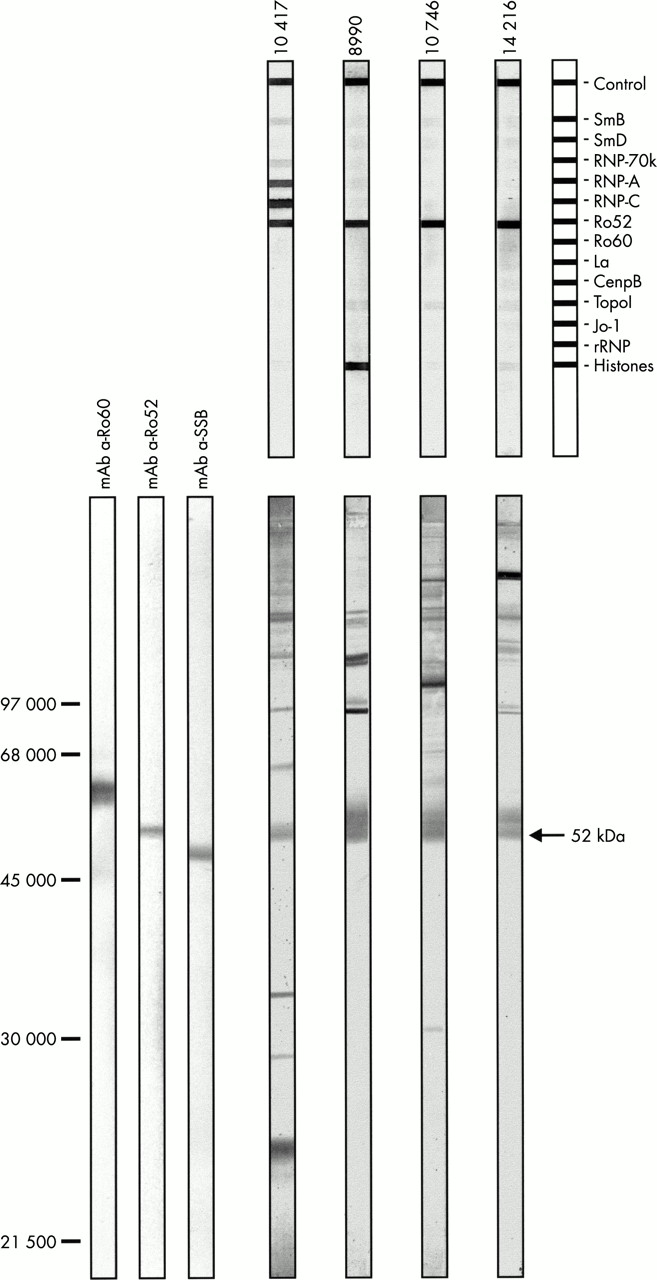
{kind=link}
{kind=link}
Confirmation of anti-Ro52 reactivity by HeLa-S100 immunoblot. Four sera reactive with Ro52 on line immunoassay (Lane 10417, 8990, 10746, 14216) (top) without detectable anti-Ro60 and anti-La/SSB antibodies. Confirmation of this anti-Ro52 reactivity by HeLa-S100 immunoblot (bottom). Monoclonal anti-Ro60 antibody (lane mAb a-Ro60). Monoclonal anti-Ro52 antibody (lane mAb a-Ro52). Monoclonal anti-SSB antibody (lane mAb a-SSB).
No serum showed reactivity in the commercial recSSA-ELISA, claimed to contain recombinant Ro52 and Ro60 in an unspecified ratio. By contrast, the SSA-ELISA with both recombinant proteins in equal concentrations confirmed nine sera. The ELISA with solely baculovirus-Ro52 confirmed three additional sera (Nos 11962, 14216, 15296). Because the Zn2+ binding domain of the Ro52 protein is a target for conformation dependent anti-Ro52 antibodies and because the antigenicity increases by reducing conditions that promote Zn2+ binding,8 we performed this ELISA under reducing and non-reducing conditions. By reducing, anti-Ro52 antibodies were found in four additional cases (Nos 8990, 10746, 12882, 16120) compared with non-reducing conditions. Thus, in 16/18 sera anti-Ro52 antibodies were confirmed by at least two other techniques (recombinant and/or natural Ro52). No nuclear or cytoplasmic staining common to the anti-Ro52 sera was identified.
Analysis of cross reactivity
Eluting antibodies from immunoblot with natural Ro60 showed no cross reactivity between these antibodies and Ro52 on line immunoassay. Preincubation with natural Ro60 of sera with both anti-Ro52 and anti-Ro60 antibodies only affected anti-Ro60 binding on line immunoassay.
Clinical correlations
In 12/18 sera, a classical connective tissue was diagnosed (table 1).
DISCUSSION
The 18 sera consecutively identified represent an additional class of anti-SSA antisera because this anti-Ro52 reactivity was not detected by conventional techniques. We believe generally used anti-SSA/Ro serological assays detect anti-Ro60 antibodies, but fail to detect additional anti-Ro52 antibodies.12 In this study the commercially available SSA-ELISA with recombinant Ro52 and Ro60 did not detect anti-Ro52 reactivity. We showed that an SSA-ELISA with recombinant Ro52 and Ro60 should contain both antigens in a 50/50 ratio to allow detection of anti-Ro52 reactivity. SSA-ELISAs based on commercial natural SSA/Ro only disclosed anti-Ro60 reactivity because this substrate does not contain detectable Ro52.13 Finally, none of our sera were positive on DID thymus/spleen. It has been shown that all sera positive on DID immunoprecipitate a metabolically labelled 60 kDa antigen.14 Immunoprecipitation in agar detects antibodies against native proteins, whereas immunoblotting preferentially detects antibodies targeting linear epitopes. Apparently exclusive anti-Ro52 antisera as defined mostly by immunoblot can be falsely negative for anti-Ro60 reactivity because this may solely be dependent on the tertiary structure of Ro60.2
Itoh et al described the anti-Ro52 immunoresponse as a cross reactive subset of the anti-Ro60 immunoresponse.15 We could not provide evidence for such a cross reactivity.
The major differences between the current series of anti-Ro52 sera and anti-Ro52 sera reported by other groups are the consecutive collection in a routine laboratory, rather than within a selected disease, the exhaustive search for and exclusion of anti-Ro60 antibodies, and the verification of anti-Ro52 by an independent assay.
We confirmed anti-Ro52 reactivity by assays using recombinant and natural Ro52. ELISA with recombinant Ro52 seemed more sensitive under reducing circumstances which prevent formation of S-S bonds and allow proper folding of Zn2+ finger domains. Controversial data on the association of Ro52 with the RoRNP particle may reflect different levels of reduced Zn2+ fingers in the preparations. 8
We found anti-Ro52 without anti-Ro60 reactivity in SLE, Sjögrens' syndrome, scleroderma, and dermatomyositis after exclusion of conformational anti-Ro60 reactivity. Rutjes et al described anti-Ro52 with anti-Jo1 antibodies,12 we observed concurrence of anti-Ro52 with anti-Cenp-B, anti-RNP, and antihistones.
In consecutive ANA positive sera, anti-SSA is present in 10.5%.1 In 88% of these anti-SSA antisera, anti-Ro52 antibodies are present (data not shown). However, anti-Ro52 without anti-Ro60 is found in approximately 1% of ANA positive sera, and constitutes an independent subset of autoreactivity. Probably, this specific profile is linked to a particular disease,12 or a specific symptom.
In conclusion, anti-Ro52 positive sera without anti-Ro60 and anti-La/SSB reactivity can be considered as an independent group, missed by classic anti-SSA/Ro detection methods: precipitin negative, not retrieved by SSA/Ro ELISAs based on natural SSA/Ro, and no specific ANA fluorescence pattern. Their expression is mostly associated with connective tissue diseases, although their precise clinical significance is under study.
Acknowledgments
We thank Caroline Dobbels for excellent assistance.
This work was supported by an IWT grant (Wetenschappelijk technologisch onderzoek in de industrie) Nos 940050 and 960267. Isabelle Peene is supported by a BOF grant from the University of Ghent.